
1. INTRODUCTION
A global agricultural boom was sparked by the emergence of the green revolution in the second half of the 20th century. The green revolution greatly increased food yields and plant productivity by bringing in new high-yielding seed varieties and using more synthetic fertilizers, insecticides, pesticides, and other agrochemicals [1,2]. Since then, there has been a significant shift in the agricultural environment worldwide. Over the past few decades, there has been a global decline in agricultural production due to the widespread usage of synthetic agrochemicals to increase crop output, which has damaged the biological and physicochemical health of the arable soil [3]. Biological wealth is being depleted and land resources are becoming smaller in the current situation. Concurrently increasing agricultural crop yield and productivity with the production of commodities related to agriculture is necessary to meet the growing need for sustainable agriculture [4]. The aforementioned complex, ecological, socioeconomic, and technical issues that arise in advancing sustainable agriculture cannot be resolved in a single or simple way [5].
An efficient way to counteract the rapid environmental degradation while maintaining high agricultural productivity and improved soil health is to promote sustainable agriculture, which gradually reduces the use of synthetic agrochemicals and increases the use of materials derived from biowaste as well as the biological and genetic potential of crop plants and microorganisms [6,7]. Certain members of the soil microbial community, especially those found in the plant rhizosphere, may help plants prevent or partially overcome environmental challenges in addition to genetically modifying the crop's physiology and metabolism to increase yield [8]. The search for environmentally appropriate substitutes to lessen the negative impacts of hazardous agrochemicals resulted in the development and application of biofertilizers, biopesticides, and other microbial-based products, such as vermicompost teas and organic extracts [9]. These microbiological products are safe for the environment, non-toxic, and have the potential to be used as instruments for disease prevention and plant growth stimulation [5].
Therefore, using microbial formulations to fertilize agricultural crops could boost the biological potential and fertility of soil while reducing the harmful impacts of agrochemicals [10]. A viable alternative to reducing the use of synthetic agrochemicals in crop production is the use of effective plant growth-promoting rhizobacteria (PGPR) as biological control agents and biofertilizers [11]. The present review concisely and holistically provides deeper insights into the various aspects of PGPR, their prospects and constraints, and their significance in encouraging the growth of plants for agriculture sustainability.
2. REGULATION OF NUTRIENT UPTAKE IN PLANTS
2.1. Nitrogen Fixation (Symbiotic and Non-symbiotic)
Plant growth-promoting (PGP) microbes have the potential to serve as a viable biological source of nitrogen fixation and its mobilization in plants, hence enhancing agricultural productivity. The minority bacterial species show the ability to fix nitrogen present in the atmosphere but are generally obligate to the most common type of symbiotic relationship within legume plants that possess a special root structure (nodules with bacteroides) harbored by colonies of bacteria. This type of symbiotic relationship is most common in diazotrophs, and the process is often termed “symbiotic biological nitrogen fixation” (e.g., Rhizobium, Bradyrhizobium, Azorhizobium, Mesorhizobium, Allorhizobium, and Sinorhizobium). This type of association can be found in many nodulating crops (peas, clover, beans, and fenugreek) Dayoub et al. [12] or in nitrogen-fixing trees (Dalbergia, Acacia, Albizia, and Leucaena) [13]. Nodules have the ability to fix atmospheric N2 via nitrogenase enzyme to ammonia. Symbiotic biological nitrogen fixation can also be found in some non-legume associations (Frankia) or stem nodulating (Azorhizobium) or leaf nodulating (Klebsiella or Burkholderia) plants. Non-nodulating pseudo-symbiotic nitrogen fixation occurs in plants and microbes, such as Spirillum, Anabaena, or Nostoc. Unlike symbiotic forms of association, there are some free associations of bacteria that fix N non-obligatory. A loose form of associative biological nitrogen fixation occurs when bacteria live between the soil or roots and use fixed N2 to exchange nutrients with plants (Fig. 1). Many other PGPR known to possess N-fixing activity include Erwinia, Acinetobacter, Alcaligenes, Pseudomonas, Ralstonia, Variovorax, Arthrobacter, Bacillus sp., and so on. [14]. A study reported that nitrogen-fixing bacteria identified as Stenotrophomonas maltophilia was improved the growth of rice plant [15]. Another study reported that, inoculation of nitrogen-fixing bacterial strains namely Azotobacter sp., Azospirillum sp., Rhizobium sp., and Pseudomonas sp. improve the growth of wheat [16]. A study concluded that inoculation of N-fixing bacterial strain identified as Erwinia rhapontici enhances the growth and physiological parameters of amaranth [17]. A similar study concluded that inoculation of Azotobacter beijerinckii improve wheat growth and soil improvement in saline-alkali land, and provides a new effective strategy for improving saline-alkali soil quality and increasing crop productivity [18]. A study by, Rana et al. [19] reported that endophytic bacterium identified as Rahnella aquatilis improves the growth of wheat crops.
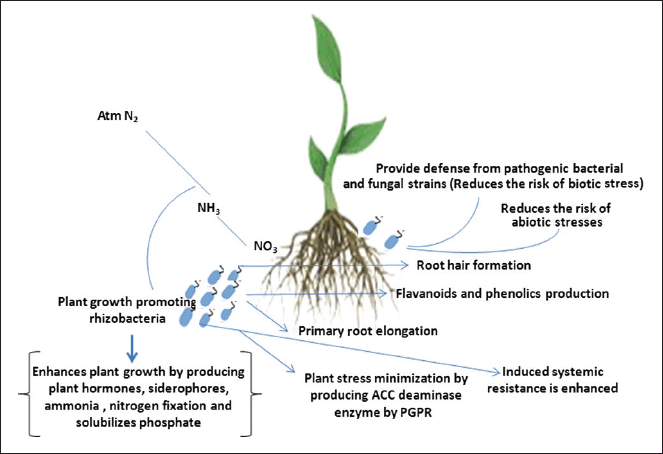 | Figure 1. The mode of action used by PGPR toward growth promotion in plants adopted from Rai et al. [30]. [Click here to view] |
2.2. Solubilization of Phosphorous
Soil phosphorous is present in organic form as the biomass of partially degraded organic matter of living organisms as in humus and other components [20]. Alternatively, it is present as inorganic complexes of insoluble Fe, Ca, Al ions, and mineral phosphates such as silicate. A fair fraction of P is non-labile to plants and this insoluble P has higher accessibility than the other reserves to plants. The inorganic form of phosphate present in the soil that is available to plants is HPO42- and H2PO4- [1]. Whereas organic phosphate (such as nucleic acids and phosphoglycerates) are inositol phosphate esters, which are dephosphorylated by the action of microbial phosphatases. Organic P is hydrolyzed as inorganic P and inositol during the phosphatase reaction [21]. Various microorganisms release this enzyme, such as Bacillus, Pseudomonas, Rhizobium, Flavobacterium, Arthrobacter Erwinia, Beijerinckia, Escherichia, Serratia, Microbacterium, Burkholderia, Staphylococcus, Micrococcus, Penicillium, Mucor, Aspergillus, Rhizopus, and Meyerozyma [22,23]. Most symbiotic, non-symbiotic, and mycorrhizal associations can solubilize P around roots. Enzymes involved in P hydrolysis include organophosphate hydrolysis enzymes (phosphomonoesterases such as phytases, phosphodiesterases, phosphotriesterases, polyphosphates, pyrophosphatase) [24–26] which are produced by microorganisms in acidic or basic soil type. Plants also produced some phosphomonoesterases from root exudates. Transformation of phosphate [inorganic P form: (acidic soil: FePO4. 2H2O, AlPO4.2H2O; alkaline soil: tricalcium phosphate, dicalcium phosphate, hydroxyapatite, and fluorapatite); organic form: (inositol phosphate, nucleotides, phospho-proteins, sugar-phosphate, phytate, and phytin)] to make it bioavailable in the form of soluble phosphate for plants [27,28]. With the secretion of organic acids from microorganisms and the release of hydroxyl or carboxyl ions, there is a reduction in pH (acidification) and a release of P around the microbial cells. The release of organic acids, i.e., succinate, gluconate, citrate, oxalate, tartrate, acetate, and so on, causes acidification. A study reported that, P- solubilizing bacterial strains identified as Pseudomonas fluorescens, Pseudomonas putida, Enterobacter sp., Bacillus megaterium, Bacillus firmus, and Pantoea agglomerans were evaluated on maize crops. The result of this study revealed that inoculation of P-solubilizing strains improves fresh and dry shoot weight, root weight, and P-nutrition uptake in mailze plant [29]. Another study revealed that, inoculation of P-solubilizing bacterial strains identified as Pseudomonas moraviensis, Bacillus halotolerans, Enterobacter hormaechei, and Pseudomonas frederiksbergensis enhances the growth of wheat crop [31]. A study concluded that, bacterial strain namely Pseudomonas azotoformans was isolated from soil in Southern Algeria, and inoculation of this strain improved seed germination percentage, shoot and root length, and fresh and dry weights as compared to uninoculated control [32]. Another study concluded that, inoculation of P-solubilizng bacteria namely Bacillus thuringiensis enhance growth and physiological parameters of sweet pepper [33]. Jiao et al. [34] reported that, inoculation of phosphorus solubilizing bacterial sp. namely Pseudomonas migulae, Pseudomonas poae, and Pseudomonas extremaustralis showed the highest soil N, P, and K content, leaf N, P, and K content and biomass as compare to control.
2.3. Potassium Solubilization
Potassium is one of the crucial nutrients required by plants in large amounts. It is present in mineral forms such as mica, feldspar, biotite, muscovite, illite, and orthoclase. PGPR plays a vital role in making avail potassium for plants [35]. Plants release different exudates that provide nutrients as well as stimulate root-colonizing bacteria and these bacteria interact with surrounding regions for nutrient sequestration. Its availability in soil is in the form of a solution, exchangeable and non-exchangeable ions, and mineral rocks. K is present as insoluble minerals rocks (silicate rocks (e.g., orthoclases), muscovite, biotite, microcline, feldspar, illite, and mica (e.g., biotite and muscovite) in soil and it is solubilized by many bacteria through organic acid action which dissolves the rock minerals, thereby increasing chelating mineral ions [36]. Chelation and ligand formation by the release of acid ions increase mineral release from rocks. Low pH increases soil acidification, enhanced chelation of cations that bound with K due to organic acid produced by bacteria provides dissolution, the release of various minerals, and the polysaccharide layer production [37,38]. Different microbes that play a role in K solubilization include Paenibacillus sp., Bacillus sp. viz B. megaterium, B. edaphicus, B. circulans, and B. mucilaginosus, Burkholderia, Pseudomonas are some PGPR that solubilizes potassium [39]. A study reported that inoculation of K-solubilizing bacterial strain identified as Klebsiella oxytoca improve maize plant height, shoot biomass, root biomass, root length, plant K uptake, grain starch, protein, oil content as well as photosynthetic rate, transpiration rate, and water use efficiency compared to control [40]. Another study reported that, bacterial strain isolated from rhizosphere of wheat and identified as Bacillus pumilus, Bacillus simplex, and B. megaterium. The study revealed that inoculation of this strain enhances growth and yield over control [41]. A study documented that, inoculation of B. megaterium improve the growth and physiological parameters of wheat plant [42]. Gandhi et al. [43] reported that bacterial strains identified as Bacillus fungorum, B. paramycoides, and Pseudomonas aeruginosa enhances growth of wheat. Kaur et al. [7] reported that inoculation of K solubilizing bacterium namely Pseudomonas gessardii (51.3 ± 1.7 mg/ml) improves the growth and physiological parameters of eggplant (Solanum melongena L.).
2.4. Phytohormone Production
PGPR can produce some phytohormones (auxin, ethylene, cytokinin, gibberellin, and abscisic acid) that are essentially needed by plants for their growth and maturity [44]. Phytohormones regulate plant physiology through chemical signaling in response to environmental changes favoring seed germination, branching, flowering, fruiting, and leaf senescence. They are required by plants in a very low concentration, i.e., less than 1 mM. PGPR produces secondary metabolites product that regulate gene expression in living organisms, vegetative, as well as reproductive growth of plants and their responses to stress, are endogenous signaling mechanisms of phytohormones [45]. Microorganisms provide these endogenous phytohormones through rhizospheric or phyllosphere interaction with plants, relieving and altering the hormone balance with variations in environmental conditions. Phytohormones produced by such rhizobacteria often activate the signaling of other hormones [viz., jasmonic acid (JA), salicylic acid (SA), brassinosteroids, and nitric oxide] available in plants in a cascade [46]. Hence, rhizobacteria can indirectly stimulate plants to adjust levels of various phytohormones that are exclusively present in their systems. Under nutrient-deprived conditions, the use of PGPR with high phytohormone production is an ideal biofertilizer in agriculture to increase the yield of different crops [47]. A study reported that inoculation of indole acetic acid-producing bacterial strain identified as Streptomyces hydrogenans DH16 significantly enhanced seed germination, shoot length, root length, fresh and dry weights, and lateral root of Pisum sativum [48]. Another study reported that inoculation of PGP bacteria identified as Priestia aryabhattai and P. frederiksbergensis produce gibberellin and IAA effectively promoted the germination of mallow and broccoli plants under saline conditions [49]. A study documented that, IAA producing Pseudomonas sp. and Sphingobium sp. were isolated from Ceanothus velutinus plant and inoculation of both of isolates improves the growth of Arabidopsis thaliana [50].
2.5. Siderophores Formation
Siderophores (Fe-celating agents) are “iron bearing” <10,000 Da low molecular weight molecules comprising ligands specific for Fe. Siderophores can be divided into catecholate type, hydroxamates type, and carboxylate type based on ligands chelating the ferric ion and several varieties of bacterial siderophores combine the various functional groups [51]. Secretion of siderophores nearby efficient PGPR ensures solubilization of iron through siderophore-iron complex formation. As Fe (III) attaches to ligands due to higher affinity, it is scavenged by these low molecular weight compounds and reduced to Fe (II) inside both microbial and plant cells. This ensures the bioavailability of iron to a wide range of bacteria and fungi that secrete siderophores [52]. After the utilization of this siderophore-Fe, the siderophore is returned to the surface of the cell. Plants utilize the advantage to take up the siderophore for their own Fe requirement [53]. Microorganisms use an active transport mechanism to recognize ferric-siderophores via membrane receptor complex. The ability of soil microorganisms to produce siderophores is vital for plant growth. A study reported that, PGP bacteria identified as Gluconacetobacter diazotrophicus and Azospirillum brasilense contribute to the iron nutrition of strawberry plants through siderophores production [54]. Another study reported that, siderophore producing bacteria namely, Pseudomonas fluorescence improve growth of Alfalfa by alleviating cadmium stress [55]. A study documented that, inoculation of siderophore producing bacterium identified as Bacillus subtilis improves growth, yield and iron content of groundnut [56]. Syed et al. [57] documented that siderophore producing Trichoderma spp. and P. fluorescence enhnace the growth, biochemical features and yield attributes of chickpea by lowering Cd uptake. A study found, efficient siderophores producer and identied as Serratia sp. EU-C1RK1 (69.16 ± 0.71 psu). The study revealed that, inoculation of this bacterium enhance growth and nutrient uptake of oats (Avena sativa L.) [6].
3. STRESS TOLERANCE MECHANISMS
3.1. 1-Aminocyclopropane-1-carboxylate (ACC) Deaminase Production
The gaseous hormone ethylene (ET) is produced by all higher plants, while some bacteria, fungi, and other species can also synthesise it. Since ethylene is a gaseous plant hormone, it can be passively diffused across plant tissues quickly and without the aid of certain transporters [58]. Even at very low concentrations, typically less than 1.0 ppm (i.e., 1.0 μl−1), ET can cause plant reactions. Even in the absence of stress, ET has been found to be active at concentrations as low as 0.05 μl/l. This is significant because ethylene is produced in a wide variety of amounts depending on the environmental conditions [59]. ET can cause seed germination, plant root elongation, development of leaf and root primordia in stems and roots, and the start of flowering at low concentrations. The enzyme S-adenosyl-methionine (SAM) synthetase, also referred to as AdoMet-synthetase, catalyses the synthesis of SAM from the combination of methionine and adenosine triphosphate, which is the first step in the synthesis of ethylene in plants. The enzyme ACC synthase then converts SAM to ACC. Furthermore, the enzyme ACC oxidase transforms the ACC into ethylene [60]. This process produces a number of other products besides ethylene, such as the volatile substances carbon dioxide and hydrogen cyanide. Plants that have been studied have been shown to have several copies of the ACC synthase and ACC oxidase genes. Several stresses, such as flooding, metal contamination of the soil, plant injuries, and elevated salt levels in the soil, cause the transcription of several of the ACC synthase genes [61].
Agrobacterium, Azospirillum, Rhizobium, Methylobacterium, Alcaligenes, Bacillus, Rhodococcus, Burkholderia, Enterobacter, Sinorhizobium, Pseudomonas, Ralstonia, and Variovorax are among the many PGP microorganisms that have been found to possess the ACC deaminase trait [62]. ACC deaminase-containing rhizobacteria have been shown to improve salt tolerance, which in turn encourages rice plant development under salt stress [63]. The inoculation of rhizobacterial strains that generated ACC deaminase improved maize growth and yield at all assessed salinity levels [64]. The application of three ACC-producing microbial consortiums, including Ochrobactrum pseudogrignonense, B. subtilis, and Pseudomonas sp., significantly increased the dry weight of the treated plants, length of the roots and shoots, and the percentage of seeds that germinated [65]. A study concluded that, ACC deaminase producing rhizobacterial strain namely Enterobacter cloacae, Serratia ficaria and Burkholderia phytofirmans were used as consortia and single-strain inoculations. The results showed that inoculation of these strain improves growth and yield of wheat (Triticum aestivum) [66]. According to a study by Khan and Singh [67] reported that, inoculation of ACC deaminase-producing Pseudomonas sp. augment drought stress tolerance and nutrient status of wheat. Another study revealed that, a salt-tolerant endophytic bacterium namely Bacillus altitudinis NKA32 with ACC deaminase activity modulates physiochemical mechanisms in rice for adaptation in the saline ecosystem [68].
3.2. Induced Systemic Response
Plants trigger a response against microorganisms irrespective of whether they are pathogenic or beneficial. Systemic acquired resistance (SAR) is related to pathogenic microorganisms, whereas induced systemic resistance is related to beneficial ones [69]. The SA mediated pathway is triggered SAR in infected sites via signaling molecules, sometimes causing necrosis in plants. While ethylene and jasmonate pathways respond to induce systemic response (ISR), helping plants overcome pathogens. In ISR, bacterial flagellar and lipoproteins, lipopolysaccharides, pyoverdine, O-side chains, and signaling molecules all play a role in the triggering mechanism [70]. Hence, PGPR used in different crops provides an indirect defense strategy against many plant pathogens.
ISR is a fascinating phenomenon orchestrated by certain PGPR that involves the activation of a plant's innate defense mechanisms against a wide spectrum of pathogens. PGPR initiates ISR through several interconnected processes. Firstly, they prime the plant, essentially preparing it to respond more effectively to potential threats. This priming involves molecular and biochemical changes that act like an immune memory system [71]. PGPR also releases signaling molecules, such as SA, JA, and ET, which serve as messengers to alert the plant of looming pathogen dangers. These signals activate various defense pathways, including SAR, a well-documented defense mechanism involving the production of antimicrobial compounds and pathogenesis-related proteins [72]. ISR also involves in the enhancement of secondary metabolites production with antimicrobial properties and the reinforcement of physical barriers like cell walls. PGPR plays a role in regulating plant hormones and improving nutrient uptake, ultimately contributing to a more robust and resilient plant defense system. Furthermore, PGPR microbial competition in the root zone can effectively outcompete pathogenic microorganisms, reducing the risk of pathogen colonization [73]. PGPR-induced ISR is a complex and multifaceted process that fortifies plants against a multitude of potential threats, fostering healthier and more resilient crops while minimizing the need for chemical pesticides.
Plant defense mechanisms that are induced by ISR include cell wall reinforcement Ahn et al. [74], the production of secondary metabolites Choudhary et al. [75], and the accumulation of defense-related enzymes (chitinases, glucanase, peroxidase, phenylalanine ammonia lyase, and polyphenol oxidase) [23]. Beneficial rhizobacteria can be used to bioprime plants, giving them systemic resistance to a variety of plant diseases, such as bacteria, viruses, and fungi. Additionally, data from a variety of fields and cannabis trials has validated PGPR's role as ISR mediators against a few insects and nematodes [76,77], hence, they can be applied for potential control. PGPR that are ISR mediators include B. subtilis, B. amyloliquefaciens, B. cereus, B. pasteurii, B. sphaericus, B. mycoides, B. pumilus, Rhizobium leguminosarum, P. fluorescence, P. putida, Serratia marcescens Bhattacharyya and Jha [23] and endophytic Actinobacteria Jacob and Sudini [78].
3.3. Microbial Volatile Organic Compounds (VOCs)
Many bacteria secrete different types of small organic compounds (molecular weight <300 g mol−1), which are small and volatile (gaseous) in nature. These molecules interact between bacteria and plants and stimulate phytohormone signaling and ISR, strengthen defense against phytopathogens, and stimulate photosynthesis. VOCs include compounds with functional groups ketones, aldehydes, terpenes, sulfides, fatty acids, and indoles. VOCs produced by microbes are effective in plant microbes signaling against pathogenic ones. These compounds exist in both solid and liquid states in soil pores, re-volatilized in organic matter or minerals surface, thus promoting their absorption by plants [79]. These VOCs are able to trigger other phytohormones within plants. VOCs assist bacteria in different roles such as the production of different biocontrol compounds, iron acquisition, phytohormones, ISR triggering, and induction of growth [80].
A study concluded that proline and chlorophyll content increased and root Na+ buildup decreased in soybeans (Glycine max) exposed to 100 mM NaCl thanks to a putative VOC mix produced from Pseudomonas simiae AU. The increase of RuBisCO long-chain proteins (photosynthesis) and vegetative storage proteins (Na+ homeostasis) in exposed soybean seedlings was validated by protein expression analysis [81]. In vitro (150 mM NaCl/15 mM CaCl2) and in soil (200 mM NaCl/20 mM CaCl2), Paraburkholderia phytofirmans PsJN VOCs have been shown to promote plant development and induce salt tolerance. The rosette area, fresh weight, and primary root length of Arabidopsis plants were all greater than those of the control plants, and exposure to VOCs demonstrated the growth-promoting effects of direct bacterial inoculation in parallel. In order to simulate the effects of VOCs, the plants were exposed to a mixture of 2-undecanone, 7-hexanol, and 3-methylbutanol molecules [82].
3.4. Biofilms
The colonized population of microorganisms around the rhizosphere coordinates their action to survive and multiply themselves around a habitat [44,83,84]. Bacteria use quorum sensing (QS) to survive different habitats by secreting signaling molecules to counteract pathogens in inhabitat plants. Rhizobacteria are capable of developing biofilms in response to Acyl-homoserine lactones [85]. QS is involved in forming dense films called biofilms; these films are frequently involved in inhabiting the neighbor territory, keeping them alive in stresses, i.e., drought, as well as low nutrients [86]. In symbiosis, the formation of biofilm is essential for root colonization and even involved in nodulation, as seen in Glycine max cv Osumi. Biofilm-active compounds such as surfactants are involved in biocontrol agents against pathogens [87]. Biofilm is present with different biomolecules like proteins, lipids, nucleic acids, and humic substances other than polysaccharides [88]. Effective colonization of roots by PGPR contributes more biofilm for layers around roots; these layers trap the nutrients and hence are beneficial during stress to plants and protect against phytopathogens [89].
3.5. Phytopathogens Biocontrol
The biocontrol of plant disease is an important characteristic of rhizobacteria. They provide host plants with benefits such as growth promotion and disease control. PGPR inhibits the growth of pathogenic bacteria, nematodes, fungi, and oomycetes via antagonism and suppresses diseases. Their proliferation and colonization steps are important in the biocontrol of the root system [90]. Biocontrol agents secreted by PGPR include 2,4-diacetyl phloroglucinol, fengycin, surfactin, phenazine, pyrrolnitrin, hydrogen cyanide, pyoluteorin, mycosubtilin, and antifungal cyclic lipopeptides [91]. Lytic enzymes, i.e., cellulases, chitinases, proteases, as well as glucanases, help bacteria in lysis and degradation of the cell wall in many fungi [92–94]. Phytopathogenic bacteria often induce virulence with increased population density, thus increasing their autoinducer secretion through QS [44,95]. This communication is interrupted by the degradation of autoinducers through enzymes produced by PGPR that help interrupt phytopathogenic QS [96] (Table 1).
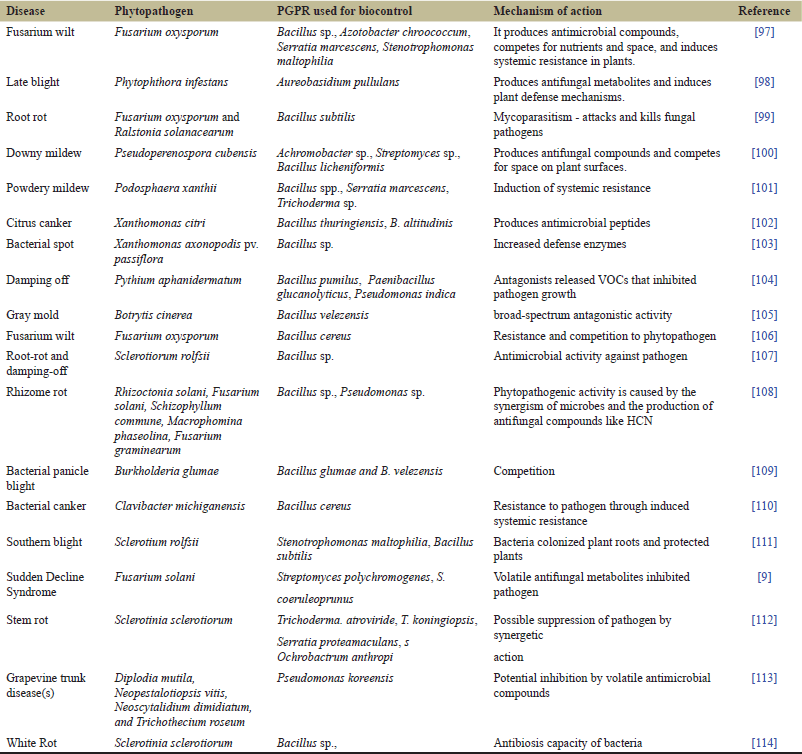 | Table 1. Mechanism and action of PGPR. [Click here to view] |
4. BIOTECHNOLOGICAL APPLICATIONS OF PGPR
4.1. Agricultural
Biofertilizers are the foundation of organic farming and they contain viable or dormant cells of efficient strains of bacteria, fungi, and algae that fix nitrogen, phosphate, potassium, and zinc, or that break down cellulosic materials [115,116]. Applying these microorganisms to seed, soil, or compost aims to boost their population and accelerate microbial activity, which increases the amount of nutrients available for plant uptake [1]. Although they do not contain any nutrients, they do help plants by releasing macronutrients and micronutrients. Furthermore, there are several direct ways that biofertilizers promote plant growth. By fixing atmospheric nitrogen both alongside and independently of plant roots, biofertilizers solubilize insoluble phosphates and other minerals, greatly increasing soil productivity. Apart from their functions, a range of bacteria and fungi can help promote plant growth by secreting hormones that directly stimulate plant growth, including gibberellins, auxin, cytokinin, abscisic acid, and indole acetic acid [117]. Plants and the majority of the beneficial microorganisms used as biofertilizers including bacteria, fungi, and cyanobacteria have symbiotic interactions. In India, a wide variety of commercial biofertilizer formulas are currently accessible. The interactions between various microorganisms and agricultural plants are being leveraged to create biofertilizers [1]. A study concluded that a nitrogen-fixing bacterial strain identified as Rahnella sp was inoculated on Aegilops kotschyi and inoculation of this bacterium improve growth and physiological parameters [118]. In an another study concluded that, endophytic nitrogen-fixing bacterium identified as R. aquatilis from cereal crops and inoculation of bacterial strain improve growth of wheat crop (Triticum aestivum L.) [19].
The infinite variety and complexity of plant diseases has led to the development of pesticides. Unfortunately, a variety of environmental problems including phytopathogen resistance have resulted from the continued usage of these herbicides [5]. Biopesticides are frequently employed to control a variety of pests, including insects and illnesses. They are produced using natural resources such as microorganisms, plants, animals, and particular minerals [119]. As of early 2013, there were over 400 registered biopesticide active ingredients and over 1,250 active biopesticide products. Sustainable organic farming operations need to use environmentally friendly pest and disease management practices along with balanced nutritional supplements to improve the quality and quantity of agricultural outputs [120]. To control pests in an eco-friendly way, a form of pesticide known as a biopesticide employs natural substances or microorganisms. It is common practice to utilize biopesticides and their byproducts to control a wide variety of pests. Their purpose is to protect crops from various pests, rodents, bacteria, and viruse [5]. Trichoderma, NPV, Bacillus thuringensis, and neem-based insecticides are the most widely developed and used biopesticides in India [121]. The quantity of microorganisms and plant and insect derivatives used as the active ingredient for the control of diseases, insects, and pests, as well as their beneficial benefits, have been reported worldwide in organic crop production. A study documented that, Paenibacillus alvei and Lysinabacillus fusiformis act as biocontrol agent against Phytophthora capsici [122]. A study revealed that, inoculation of S. marcescens inhibiting the growth of Pythium aphanidermatum [123].
4.2. Environmental
Soil metal contamination is a major problem because of the increasing number of impacted locations worldwide as well as the detrimental consequences that metals have on the environment and human health. Because metals are harmful and non-biodegradable, it is imperative to clean up the hundreds of contaminated sites worldwide and stop more metal pollution [124]. Traditional restoration methods, which rely on physical and chemical methods, are often expensive, impractical, and cause secondary environmental issues. In light of this, microbe-aided phytoremediation has grown in popularity due to its low cost, lack of negative environmental effects, and significant recent developments [125]. Microorganisms are essential to the recovery of natural ecosystems and are a fundamental part of them. Actually, for plants to survive metal toxicity and thrive in these harsh environments, plant-microbe interactions in metal-contaminated soil are crucial. Therefore, improving our understanding of this intricate relationship is crucial to the advancement of phytoremediation. Microbes are therefore crucial to the restoration of function and biodiversity in an ecosystem.
The ability of the microbial cells to create and detect signal molecules allows them to spread out as a biofilm across the root surface and begin functioning in unison once a particular population density is attained [85]. QS is the name given to this particular phenomenon. Microbes are very beneficial, essential for plant nutrition, and able to mitigate the negative effects of metals. Certain rhizosphere bacteria are able to directly handle organic and inorganic contaminants by taking advantage of their natural degrading processes, such as volatilization, transformation, and rhizo-degradation [126]. Metal complexation, EPS sequestration, volatilization, efflux, impermeability to metals, Hg(II) reduction to Hg(0), and enzymatic detoxification discharged from cells are some of the resistance mechanisms displayed by bacteria [127]. Additionally, bacterial plasmids contain resistance genes for a number of toxic-heavy metals and metalloids. PGP microorganisms can lessen a number of limiting factors for phytoremediation technologies, such as metal solubility, pollution level, and soil chemistry. A study revealed that, Trichoderma harzianum have showing tolerance in Cd-polluted soil and enhanced growth of barley [128]. Another study revelaed that, combine application of Bacillus mycoides and rock phosphate enhance growth of wheat by alleviating heavy metal stress [129].
5. COMMERCIAL APPLICATION OF PGPR
Marketing and commercialization are PGPR primary and most important tasks for success. Before entering the farming field, meticulous management techniques, adequate study, market analysis, and survey, are necessary to produce the right microbial inoculation in a laboratory. The particular crop and the climate at the time of commercial availability determine which microbial isolate is best [130]. For example, if a certain crop is susceptible to fungi, the selection process may be based on the microorganisms' particular antagonistic activity against the fungus. The performance of microorganisms is significantly influenced by the climatic conditions of their surroundings. Microbes in the laboratory are exposed to various biotic and abiotic stimuli in order to force them to combat natural environmental conditions, taking into account the climatic setting [131]. A nation's ecological zone combines the soil, climate, and temperature characteristics of a particular area. The process of creating novel microbial consortia for various ecological zones based on different crops is quite intricate and challenging. Climate factors such as rising temperatures may cause severe drought, which would impact PGPR performance in addition to crop growth and development [132]. Since native microorganisms are not naturally formed from locally rhizospheric soils, their performance has not yet been determined, and as a result, commercial inoculation may not be successful. Climate conditions can alter a plant's physiology and disrupt root exudation, which can lead to alterations in the plant's growth and development [133].
In terms of their desirable traits and colonization capabilities in specific stressful environments, PGPR may be well-known for its positive effects on plant growth and development. Understanding the disease control mechanism, growth promotion features, and resistance to biotic and abiotic variables can help predict the performance of microbial inoculants in different climatic situations [134,135]. The identification and isolation of niche-specific microorganisms with superior microbial features is required to attain maximal microbial efficiency. The needs of the crop are given high consideration while marketing microbial inoculation. Therefore, no single microbial strain is capable of performing well for every crop and in every climate. The primary and most important processes in commercialization include field surveying farmers' fields, isolating particular microbial strains with particular characteristics, choosing an appropriate carrier material, and injection technique [72].
Furthermore, commercial businesses favor microorganisms that have no negative effects on the environment or people. The laws governing the selection and screening process for commercial marketing should be regulated by the government [136]. Neglected non-targeted consequences, such as the replacement of natural species, could be another significant problem. Hypovirulent strains may become pathogenic, poisonous, or allergic. Before the widespread acceptance, registration, and implementation of PGPR for the management of pests and diseases, the aforementioned safety concerns must be addressed [137].
6. ROLE OF PGPR IN AGRICULTURE: PROMISE VERSUS BOTTLENECKS
The rising expense of agrochemicals and societal need for green technology is driving up demand for microbial inoculants. There has been a reported 12% annual growth in the global market for biostimulants [138]. Some PGPB, such as Burkholderia, Pseudomonas, Rhizobium, Azospirillum, Azotobacter, Bacillus, and Serratia sp., have been used to produce commercial products on a large scale [139]. However, different nations have varied laws governing the use of microbial inoculants in agricultural techniques [140]. The shelf-life, dependability, and uniformity of microbial inoculants in agricultural settings are the primary constraints. The shelf life of gram-negative bacteria is less than that of spore-forming gram-positive bacteria. It was suggested that super-inoculants with all the desired characteristics be used [141]. According to reports, some of the PGPB are opportunistic human pathogens such Burkholderia cepacia and Pseudomonas aeruginosa [142,143], which provide dangers to humans and the environment that should be appropriately addressed prior to their commercial production. Enhancements are required for the creation of novel nanoparticle-based carriers, more effective PGP rhizobacterial consortia, and application device optimization. Further problem is that plants are home to a variety of human infections, many of which have been shown positive effects on plant health by boosting plant growth [144–146]. More research is needed to address the issue of possibly harmful microorganisms in sustainable agriculture. The rhizosphere of plants is designed to make it easier for beneficial microorganisms to colonize than for harmful ones [147]. The biosafety of PGPB products is now being reevaluated in the United States, Europe, and other nations. Through changes in rhizosphere biology, resource availability, and biogeochemical cycling, climate change can influence the relationship between plants and microbes [148]. Once the shortcomings pertaining to long-term impacts on soil microbial populations, farmer acceptance, economic sustainability, and governmental regulations are resolved, PGPB's full potential will be achieved.
7. CONCLUSION
In addition to being essential to a country's survival, the agriculture sector helps it meet the demands of its expanding population and generate revenue for exports. The agroindustry has seen a number of technological advancements since the green revolution, which has improved crop yield but also caused environmental issues. The rising use of biological inoculants rather than agrochemicals for sustainable agriculture worldwide during the past ten years has unavoidably brought about a revolution. Maintaining the integrity of our planet's health and appropriate biogeochemical cycling, as well as the general growth and increased productivity of crop plants, depends on the triad of interactions between the bioinoculant microorganism, resident soil microbiota, and host plant. Growing concerns about food safety and the necessity to regulate the quality of food production to meet shifting customer demands are predicted to cause farmers to turn to organic farming and embrace sustainable agricultural methods. In order to find environmentally suitable substitutes for harmful chemicals, it is necessary to take into account the three important "Ps": people, prosperity, and the planet. To achieve the intended results and win over the confidence of farmers, who are the true stakeholders in agriculture, this microbial product-based technology must first undergo extensive research and development. Authentication, strain enhancement, and commercial production quantification are the main research issues that require further attention. As environmentally sustainable crop development alternatives, governments and federal agencies ought to encourage the use of biofertilizers and biopesticides. Entrepreneurs ought to contribute more to the biofertilizer sector and support new businesses financially. Furthermore, widespread public education is necessary to inform farmers and consumers alike of the benefits of employing microbe-based biopesticides and fertilizers in order to ensure a more environmentally friendly future.
8. AUTHOR CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
9. ETHICS APPROVAL AND CONSENT TO PARTICIPATE
This study does not involve experiments on animals or human subjects.
10. FUNDING
There is no funding to report.
11. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
12. PUBLISHER’S NOTE:
All claims expressed in this article are solely those of the authors and do not necessarily represent those of the publisher, the editors and the reviewers. This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
13. USE OF ARTIFICIAL INTELLIGENCE (AI)-ASSISTED TECHNOLOGY
The authors declares that they have not used artificial intelligence (AI)-tools for writing and editing of the manuscript, and no images were manipulated using AI.
14. DATA AVAILABILITY
All the data is available with the authors and shall be provided upon request.
REFERENCES
1. Negi R, Sharma B, Jan T, Kaur T, Chowdhury S, Kapoor M, et al. Microbial consortia: promising tool as plant bioinoculants for agricultural sustainability. Curr Microbiol 2024;81:222; CrossRef
2. Sharma S, Chaitanya MVNL, Sharma S, Kumar S, Rustagi S, Singh S, et al. The medicinal plant Berberis aristata and its endophytes for pharmacological applications: current research and future challenges. J Appl Biol Biotechnol 2024;12:37–46. CrossRef
3. Ahemad M, Kibret M. Mechanisms and applications of plant growth promoting rhizobacteria: current perspective. J King Saud Univ Sci 2014;26:1–20. CrossRef
4. Rachappanavar V, Kumar M, Negi N, Chowdhury S, Kapoor M, Singh S, et al. Silicon derived benefits to combat biotic and abiotic stresses in fruit crops: current research and future challenges. Plant Physiol Biochem 2024;211:108680; CrossRef
5. Negi R, Sharma B, Kaur S, Kaur T, Khan SS, Kumar S, et al. Microbial antagonists: diversity, formulation and applications for management of pest–pathogens. Egypt J Biol Pest Control 2023;33:105; CrossRef
6. Devi R, Alsaffar MF, Al-Taey DKA, Kumar S, Negi R, Sharma B, et al. Synergistic effect of minerals solubilizing and siderophores producing bacteria as different microbial consortium for growth and nutrient uptake of oats (Avena sativa L.). Vegetos 2024;37:1863–75; CrossRef
7. Kaur T, Devi R, Negi R, Kumar S, Singh S, Rustagi S, et al. Microbial consortium with multifunctional attributes for the plant growth of eggplant (Solanum melongena L.). Folia Microbiol 2024;69:1255–66; CrossRef
8. Sonowal T, Gupta N, Kumar S, Rustagi S, Singh S, Rai AK, et al. Plant growth-promoting rhizobacteria: influence to abiotic stress tolerance in rice (Oryza sativa L.). J Appl Biol Biotechnol 2024;12:41–7. CrossRef
9. Alblooshi AA, Purayil GP, Saeed EE, Ramadan GA, Tariq S, Altaee AS, et al. Biocontrol potential of endophytic actinobacteria against Fusarium solani, the causal agent of sudden decline syndrome on date palm in the UAE. J Fungi 2021;8:8; CrossRef
10. Choudhary M, Meena VS, Yadav RP, Parihar M, Pattanayak A, Panday S, et al. Does PGPR and mycorrhizae enhance nutrient use efficiency and efficacy in relation to crop productivity? In: Maheshwari D, Dheeman S (eds.). Field crops: sustainable management by PGPR. Sustainable development and biodiversity, Springer, Cham, Switerzland, pp 45–68, 2019; CrossRef
11. Grover M, Bodhankar S, Sharma A, Sharma P, Singh J, Nain L. PGPR mediated alterations in root traits: way toward sustainable crop production. Front Sustain Food Syst 2021;4:618230. CrossRef
12. Dayoub E, Naudin C, Piva G, Shirtliffe SJ, Fustec J, Corre-Hellou G. Traits affecting early season nitrogen uptake in nine legume species. Heliyon 2017;3:e00244. CrossRef
13. Dhiman VK, Rana N, Dhiman VK, Pandey H, Verma P, Singh D. Effect of rhizobial isolates and nitrogen fertilizers on nursery performance, nodulation behavior and nitrogenase activity of Dalbergia sissoo Roxb. seedlings. Plant Stress 2022;4:100080. CrossRef
14. Fuentes-Ramirez LE, Caballero-Mellado J. Bacterial biofertilizers. In: Siddiqui ZA (ed.). PGPR: biocontrol and biofertilization, Springer, Dordrecht, The Netherlands, pp 143–72, 2006; CrossRef
15. Swarnalakshmi K, Yadav V, Tyagi D, Dhar DW, Kannepalli A, Kumar S. Significance of plant growth promoting rhizobacteria in grain legumes: growth promotion and crop production. Plants 2020;9:1596. CrossRef
16. Din I, Khan H, Khan NA, Khil A. Inoculation of nitrogen fixing bacteria in conjugation with integrated nitrogen sources induced changes in phenology, growth, nitrogen assimilation and productivity of wheat crop. J Saudi Soc Agric Sci 2021;20:459–66. CrossRef
17. Devi R, Kaur T, Kour D, Yadav AN. Microbial consortium of mineral solubilizing and nitrogen fixing bacteria for plant growth promotion of amaranth (Amaranthus hypochondrius L.). Biocatal Agric Biotechnol 2022;43:102404. CrossRef
18. Zhou L, Liu W, Duan H, Dong H, Li J, Zhang S, et al. Improved effects of combined application of nitrogen-fixing bacteria Azotobacter beijerinckii and microalgae Chlorella pyrenoidosa on wheat growth and saline-alkali soil quality. Chemosphere 2023;313:137409. CrossRef
19. Rana KL, Negi R, Sharma B, Yadav A, Devi R, Kaur T, et al. Potential effect of novel endophytic nitrogen fixing diverse species of Rahnella on growth promotion of wheat (Triticum aestivum L.). J Crop Sci Biotechnol 2024;27:605–15; CrossRef
20. Biswas T, Kole SC. Soil organic matter and microbial role in plant productivity and soil fertility. In: Adhya TK, Mishra BB, Annapurna K, Verma DK, Kumar U (eds.). Advances in soil microbiology: recent trends and future prospects: volume 2: soil-microbe-plant interaction, Springer, Singapore, pp 219–38, 2017; CrossRef
21. Jarosch KA, Kandeler E, Frossard E, Bünemann EK. Is the enzymatic hydrolysis of soil organic phosphorus compounds limited by enzyme or substrate availability? Soil Biol Biochem 2019;139:107628; CrossRef
22. Alori ET, Glick BR, Babalola OO. Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front Microbiol 2017;8:971. CrossRef
23. Bhattacharyya PN, Jha DK. Plant growth-promoting rhizobacteria (PGPR): emergence in agriculture. World J Microbiol Biotechnol 2012;28:1327–50.
24. Nannipieri P, Giagnoni L, Landi L, Renella G. Role of phosphatase enzymes in soil. In: Bünemann E, Oberson A, Frossard E (eds.). Phosphorus in action. Soil biology, Springer, Berlin, Heidelberg, Germany, vol. 26, pp. 215–43, 2011; CrossRef
25. Bünemann EK. Assessment of gross and net mineralization rates of soil organic phosphorus–a review. Soil Biol Biochem 2015;89:82–98. CrossRef
26. De Castro AA, Prandi IG, Kuca K, Ramalho TC. Organophosphorus degrading enzymes: molecular basis and perspectives for enzymatic bioremediation of agrochemicals. Cienc Agrotec 2017;1:41–82. CrossRef
27. Kaur R, Kaur S. Biological alternates to synthetic fertilizers: efficiency and future scopes. Indian J Agric Res 2018;52:587–95. CrossRef
28. Mitter EK, Tosi M, Obregón D, Dunfield KE, Germida JJ. Rethinking crop nutrition in times of modern microbiology: innovative biofertilizer technologies. Front Sustain Food Syst 2021;5:606815. CrossRef
29. Sarikhani MR, Aliasgharzad N, Khoshru B. P solubilizing potential of some plant growth promoting bacteria used as ingredient in phosphatic biofertilizers with emphasis on growth promotion of Zea mays L. Geomicrobiol J 2020;37:327–35. CrossRef
30. Rai PK, Singh M, Anand K, Saurabh S, Kaur T, Kour D, et al. Role and potential applications of plant growth-promoting rhizobacteria for sustainable agriculture. In: Rastegari AA, Yadav AN, Yadav N (eds.). New and future developments in microbial biotechnology and bioengineering, Elsevier, Netherlands, The Netherlands, pp 49–60, 2020; CrossRef
31. Fahsi N, Mahdi I, Mesfioui A, Biskri L, Allaoui A. Plant growth-promoting rhizobacteria isolated from the jujube (Ziziphus lotus) plant enhance wheat growth, Zn uptake, and heavy metal tolerance. Agriculture 2021;11:316. CrossRef
32. Belkebla N, Bessai SA, Melo J, Caeiro MF, Cruz C, Nabti EH. Restoration of Triticum aestivum growth under salt stress by phosphate-solubilizing bacterium isolated from Southern Algeria. Agronomy 2022;12:2050. CrossRef
33. Devi R, Kaur T, Negi R, Kour D, Chaubey KK, Yadav AN. Indigenous plant growth-promoting rhizospheric and endophytic bacteria as liquid bioinoculants for growth of sweet pepper (Capsicum annuum L.). Biologia 2023;78:2623–33; CrossRef
34. Jiao H, Wang R, Qin W, Yang J. Screening of rhizosphere nitrogen fixing, phosphorus and potassium solubilizing bacteria of Malus sieversii (Ldb.) Roem. and the effect on apple growth. J Plant Physiol 2024;292:154142. CrossRef
35. Devi R, Kaur T, Negi R, Sharma B, Chowdhury S, Kapoor M, et al. Biodiversity, mechanisms, and potential biotechnological applications of minerals solubilizing extremophilic microbes: a review. J Appl Biol Biotechnol 2024;12:23–40. CrossRef
36. Etesami H, Emami S, Alikhani HA. Potassium solubilizing bacteria (KSB): Mechanisms, promotion of plant growth, and future prospects a review. J Soil Sci Plant Nutr 2017;17:897–911. CrossRef
37. Das I, Pradhan M. Potassium-solubilizing microorganisms and their role in enhancing soil fertility and health. In: Meena V, Maurya B, Verma J, Meena R (eds.). Potassium solubilizing microorganisms for sustainable agriculture, Springer, New Delhi, India, pp 281–91, 2016; CrossRef
38. Jaiswal DK, Verma JP, Prakash S, Meena VS, Meena RS. Potassium as an important plant nutrient in sustainable agriculture: a state of the art. In: Meena V, Maurya B, Verma J, Meena R (eds.). Potassium solubilizing microorganisms for sustainable agriculture, Springer, New Delhi, India, pp 21–9, 2016; CrossRef
39. Bhattacharjya S, Das S, Amat D. Potential of microbial inoculants for organic waste decomposition and decontamination. In: Rakshit A, Meena VS, Parihar M, Singh HB, Singh AK (eds.). Advance in bioinoculants, Elsevier, Woodhead Publishing, Langford Lane, UK, pp 103–32, 2021; CrossRef
40. Imran M, Shahzad SM, Arif MS, Yasmeen T, Ali B, Tanveer A. Inoculation of potassium solubilizing bacteria with different potassium fertilization sources mediates maize growth and productivity. Pak J Agric Sci 2020;57:1045–55.
41. Chandra P, Khobra R, Sundha P, Sharma RK, Jasrotia P, Chandra A, et al. Plant growth promoting Bacillus-based bio formulations improve wheat rhizosphere biological activity, nutrient uptake and growth of the plant. Acta Physiol Plant 2021;43:1–12. CrossRef
42. Negi R, Kaur T, Devi R, Kour D, Yadav AN. Assessment of nitrogen-fixing endophytic and mineral solubilizing rhizospheric bacteria as multifunctional microbial consortium for growth promotion of wheat and wild wheat relative Aegilops kotschyi. Heliyon 2022;8:e12579; CrossRef
43. Gandhi R, Prittesh P, Jinal HN, Chavan SM, Paul D, Amaresan N. Evaluation of the effect of potassium solubilizing bacterial strains on the growth of wheat (Triticum aestivum L.). J Plant Nut 2023;46:1479–90. CrossRef
44. Singh D, Agarwal V. Herbal antibacterial remedy against upper respiratory infection causing bacteria and in vivo safety analysis. Vegetos 2022;35:264–68. CrossRef
45. Singh D, Pandey H, Thakur K, Sharma I, Naraian R. Aquaporins and their functions in water transportation in different plant species. Environ Sustain 2022;5:443–56. CrossRef
46. Fahad S, Hussain S, Bano A, Saud S, Hassan S, Shan D, et al. Potential role of phytohormones and plant growth-promoting rhizobacteria in abiotic stresses: consequences for changing environment. Environ Sci Pollut Res 2015;22:4907–21. CrossRef
47. Amara U, Khalid R, Hayat R. Soil bacteria and phytohormones for sustainable crop production. In: Maheshwari D (ed.). Bacterial metabolites in sustainable agroecosystem. Sustainable development and biodiversity. Springer, Cham, Switerzland, pp 87–103, 2015; CrossRef
48. Kaur T, Manhas RK. Evaluation of ACC deaminase and indole acetic acid production by Streptomyces hydrogenans DH16 and its effect on plant growth promotion. Biocatal Agric Biotechnol 2022;42:102321. CrossRef
49. Woo JI, Injamum-Ul-Hoque M, Zainurin N, Shaffique S, Kwon EH, Gam HJ, et al. Gibberellin producing bacteria isolated from coastal soil enhance seed germination of mallow and broccoli plants under saline conditions. BioTech 2023;12:66. CrossRef
50. Ganesh J, Hewitt K, Devkota AR, Wilson T, Kaundal A. IAA-producing plant growth promoting rhizobacteria from Ceanothus velutinus enhance cutting propagation efficiency and Arabidopsis biomass. Front Plant Sci 2024;15:1374877. CrossRef
51. Thakur K, Pandey H, Sharma I, Dhiman V, Pandey D. 13 siderophores and their applications in heavy metal detoxification. In: Thakur K, Pandey H, Sharma I, Dhiman VK, Pandey D (eds.). Plant-microbial interactions and smart agricultural biotechnology, CRC Press, Boca Raton, FL, pp 1–298, 2021; CrossRef
52. Negi S, Singh D, Pandey H, Singhal RK. Biofortification for high Fe and Zn in various Poaceae crops by using different molecular breeding and biotechnological approaches. Plant Physiol Rep 2021;26:636–46. CrossRef
53. Ahmed E, Holmström SJ. Siderophores in environmental research: roles and applications. Microb Biotechnol 2014;7:196–208. CrossRef
54. Delaporte-Quintana P, Lovaisa NC, Rapisarda VA, Pedraza RO. The plant growth promoting bacteria Gluconacetobacter diazotrophicus and Azospirillum brasilense contribute to the iron nutrition of strawberry plants through siderophores production. Plant Growth Regul 2020;91:185–99; CrossRef
55. Sepehri M, Khatabi B. Combination of siderophore-producing bacteria and Piriformospora indica provides an efficient approach to improve cadmium tolerance in alfalfa. Microb Ecol 2021;81:717–30. CrossRef
56. Sarwar S, Khaliq A, Yousra M, Sultan T. Iron biofortification potential of siderophore producing rhizobacterial strains for improving growth, yield and iron contents of groundnut. J Plant Nutr 2022;45:2332–47. CrossRef
57. Syed A, Elgorban AM, Bahkali AH, Eswaramoorthy R, Iqbal RK, Danish S. Metal-tolerant and siderophore producing Pseudomonas fluorescence and Trichoderma spp. improved the growth, biochemical features and yield attributes of chickpea by lowering Cd uptake. Sci Rep 2023;13:4471. CrossRef
58. Kashyap P, Shirkot P, Das R, Pandey H, Singh D. Biosynthesis and characterization of copper nanoparticles from Stenotrophomonas maltophilia and its effect on plant pathogens and pesticide degradation. J Agric Food Res 2023;13:100654. CrossRef
59. Sharma A, Pandey H, Manpoong C, Vashishth A, Singh D, Bang NTH. Biometric analysis, biosynthetic pathway and multipurpose uses of buckwheat local varieties in Namsai district of eastern Himalayas in India. Trends Food Sci Technol 2023;136:251–67; CrossRef
60. Polko JK, Kieber JJ. 1-aminocyclopropane 1-carboxylic acid and its emerging role as an ethylene-independent growth regulator. Front Plant Sci 2019;10:1602. CrossRef
61. Van de Poel B, Van Der Straeten D. 1-aminocyclopropane-1-carboxylic acid (ACC) in plants: more than just the precursor of ethylene! Front Plant Sci 2014;5:640. CrossRef
62. Arshad M, Saleem M, Hussain S. Perspectives of bacterial ACC deaminase in phytoremediation. Trends Biotechnol 2007;25:356–62. CrossRef
63. Bal HB, Nayak L, Das S, Adhya TK. Isolation of ACC deaminase producing PGPR from rice rhizosphere and evaluating their plant growth promoting activity under salt stress. Plant Soil 2013;366:93–105; CrossRef
64. Nadeem SM, Zahir ZA, Naveed M, Arshad M. Preliminary investigations on inducing salt tolerance in maize through inoculation with rhizobacteria containing ACC deaminase activity. Can J Microbiol 2007;53:1141–49; %m18026206 CrossRef
65. Saikia J, Sarma RK, Dhandia R, Yadav A, Bharali R, Gupta VK, et al. Alleviation of drought stress in pulse crops with ACC deaminase producing rhizobacteria isolated from acidic soil of Northeast India. Sci Rep 2018;8:3560; CrossRef
66. Zafar-ul-Hye M, Zahra MB, Danish S, Abbas M, Rehim A, Akbar MN, et al. Multi-strain inoculation with pgpr producing acc deaminase is more effective than single-strain inoculation to improve wheat (Triticum aestivum) growth and yield. Phyton 2020;89:405–13. CrossRef
67. Khan A, Singh AV. Multifarious effect of ACC deaminase and EPS producing Pseudomonas sp. and Serratia marcescens to augment drought stress tolerance and nutrient status of wheat. World J Microbiol Biotechnol 2021;37:198; CrossRef
68. Bharti C, Fatima T, Mishra P, Verma P, Bhattacharya A, Alaylar B, et al. Salt-tolerant endophytic Bacillus altitudinis NKA32 with ACC deaminase activity modulates physiochemical mechanisms in rice for adaptation in saline ecosystem. Environ Sustain 2024;7:231–49; CrossRef
69. Yu Y, Gui Y, Li Z, Jiang C, Guo J, Niu D. Induced systemic resistance for improving plant immunity by beneficial microbes. Plants 2022;11:386. CrossRef
70. Annapurna K, Kumar A, Kumar LV, Govindasamy V, Bose P, Ramadoss D. PGPR-induced systemic resistance (ISR) in plant disease management. In: Maheshwari D (ed.). Bacteria in agrobiology: disease management. Springer, Berlin, Heidelberg, Germany, pp. 405–25, 2013; CrossRef
71. Rosier A, Medeiros FHV, Bais HP. Defining plant growth promoting rhizobacteria molecular and biochemical networks in beneficial plant-microbe interactions. Plant Soil 2018;428:35–55; CrossRef
72. Negi R, Sharma B, Parastesh F, Kaur S, Khan SS, Kour D, et al. Microbial consortia mediated regulation of plant defense: a promising tool for sustaining crops protection. Physiol Mol Plant Pathol.2024;134:102393; CrossRef
73. Kumar M, Karthikeyan N, Prasanna R. Priming of plant defense and plant growth in disease-challenged crops using microbial consortia. In: Choudhary DK, Varma A (eds.). Microbial-mediated induced systemic resistance in plant, Springer Nature, Singapore, pp 39–56, 2016; CrossRef
74. Ahn IP, Lee SW, Suh SC. Rhizobacteria-induced priming in Arabidopsis is dependent on ethylene, jasmonic acid, and NPR1. Mol Plant-Microbe Interact 2007;20:759–68. CrossRef
75. Choudhary DK, Prakash A, Johri B. Induced systemic resistance (ISR) in plants: mechanism of action. Indian J Microbiol 2007;47:289–97. CrossRef
76. Pineda A, Zheng SJ, van Loon JJ, Pieterse CM, Dicke M. Helping plants to deal with insects: the role of beneficial soil-borne microbes. Trends Plant Sci 2010;15:507–14. CrossRef
77. Singh S, Pathak R, Choudhary V. Plant growth-promoting rhizobacteria-mediated acquired systemic resistance in plants against pests and diseases. In: Choudhary DK, Varma A (eds.). Microbial-mediated induced systemic resistance in plant, Springer Nature, Singapore, pp 125–34, 2016; CrossRef
78. Jacob S, Sudini HK. Indirect plant growth promotion in grain legumes: role of actinobacteria. In: Subramaniam G, Arumugam S, Rajendran V (eds.). Plant growth promoting actinobacteria. Springer, Singapore, pp 17–32, 2016; CrossRef
79. Santoro M, Cappellari L, Giordano W, Banchio E. Production of volatile organic compounds in PGPR. In: Cassán F, Okon Y, Creus C (eds.). Handbook for Azospirillum: technical issues and protocols, Springer, Cham, Switzerland, pp 307–17, 2015. CrossRef
80. Morales-García YE, Baez A, Quintero-Hernández V, Molina-Romero D, Rivera-Urbalejo AP, Pazos-Rojas LA, et al. Bacterial mixtures, the future generation of inoculants for sustainable crop production. In: Maheshwari D, Dheeman S (eds.). Field crops: sustainable management by PGPR. Sustainable development and biodiversity, Springer, Cham, Switzerland, pp 11–44, 2019; CrossRef
81. Vaishnav A, Kumari S, Jain S, Varma A, Choudhary DK. Putative bacterial volatile-mediated growth in soybean (Glycine max L. Merrill) and expression of induced proteins under salt stress. J Appl Microbiol 2015;119:539–51; CrossRef
82. Ledger T, Rojas S, Timmermann T, Pinedo I, Poupin MJ, Garrido T, et al. Volatile-mediated effects predominate in Paraburkholderia phytofirmans growth promotion and salt stress tolerance of Arabidopsis thaliana. Front Microbiol 2016;7:1838; CrossRef
83. Singh D, Agarwal V. Screening of antimicrobial, anti-quorum sensing activity and cytotoxicity of origanum oil against gram-positive AND GRAM-negative bacteria. Biomedicine 2021;41:599–603. CrossRef
84. Singh D, Sharma D, Agarwal V. Screening of anti-microbial, anti-biofilm activity, and cytotoxicity analysis of a designed polyherbal formulation against shigellosis. J Ayurveda Integr Med 2021;12:601–6. CrossRef
85. Dhiman S, Kumar A, Kaur G, Mukherjee G, Rustagi S, Shreaz S, et al. Bacterial biofilms: pathogenesis, monitoring, treatment approaches and associated challenges. Biologia 2024;79:3161–81; CrossRef
86. Altaf MM, Khan MSA, Abulreesh HH, Ahmad I. Quorum sensing in plant growth-promoting rhizobacteria and its impact on plant-microbe interaction. In: Singh D, Singh H, Prabha R (eds.). Plant-microbe interactions in agro-ecological perspectives: volume 1: fundamental mechanisms, methods and functions, Springer, Singapore, pp 11–31, 2017; CrossRef
87. Thérien M, Kiesewalter HT, Auria E, Charron-Lamoureux V, Wibowo M, Maróti G, et al. Surfactin production is not essential for pellicle and root-associated biofilm development of Bacillus subtilis. Biofilm 2020;2:100021. CrossRef
88. Vu B, Chen M, Crawford RJ, Ivanova EP. Bacterial extracellular polysaccharides involved in biofilm formation. Molecules 2009;14:2535–54. CrossRef
89. Gupta G, Snehi SK, Singh V. Role of PGPR in biofilm formations and its importance in plant health. In: Iqbal A, Fohad MH (eds.). Biofilms in plant and soil health, Wiley, Hoboken, NJ, pp 27–42, 2017; CrossRef
90. Beneduzi A, Ambrosini A, Passaglia LM. Plant growth-promoting rhizobacteria (PGPR): their potential as antagonists and biocontrol agents. Genet Mol Biol 2012;35:1044–51. CrossRef
91. Wang H, Liu R, You MP, Barbetti MJ, Chen Y. Pathogen biocontrol using plant growth-promoting bacteria (PGPR): role of bacterial diversity. Microorganisms 2021;9:1988; CrossRef
92. Dhiman VK, Chauhan V, Kanwar SS, Singh D, Pandey H. Purification and characterization of actinidin from Actinidia deliciosa and its utilization in inactivation of α-amylase. Bull Natl Res Cent 2021;45:1–9. CrossRef
93. Chauhan V, Kaushal D, Dhiman VK, Kanwar SS, Singh D, Dhiman VK, et al. An insight in developing carrier-free immobilized enzymes. Front Bioeng Biotechnol 2022;10:794411. CrossRef
94. Sharma SK, Singh D, Pandey H, Jatav RB, Singh V, Pandey D. An overview of roles of enzymatic and nonenzymatic antioxidants in plant. In: Aftab T, Hakeem KR (eds.). Antioxidant defense in plants, Springer, Singapore, pp 1–13, 2022; CrossRef
95. Singh D, Singh V, Mishra SB, Sharma D, Agarwal V. Evaluation of anti-biofilm, anti-quorum, anti-dysenteric potential of designed polyherbal formulation: in vitro and in vivo study. J Appl Biomed 2022;20:7–17. CrossRef
96. Helman Y, Chernin L. Silencing the mob: disrupting quorum sensing as a means to fight plant disease. Mol Plant Pathol 2015;16:316–29. CrossRef
97. Aydi-Ben-Abdallah R, Jabnoun-Khiareddine H, Daami-Remadi M. Fusarium wilt biocontrol and tomato growth stimulation, using endophytic bacteria naturally associated with Solanum sodomaeum and S. bonariense plants. Egypt J Biol Pest Cont 2020;30:1–13. CrossRef
98. Di Francesco A, Milella F, Mari M, Roberti R. A preliminary investigation into Aureobasidium pullulans as a potential biocontrol agent against Phytophthora infestans of tomato. Biol Cont 2017;114:144–49. CrossRef
99. Qiao J, Zhang R, Liu Y, Liu Y. Evaluation of the biocontrol efficiency of Bacillus subtilis wettable powder on pepper root rot caused by Fusarium solani. Pathogens 2023;12:225. CrossRef
100. Konineeka S, Chandan S, Jayanta S. PGPR consortium in alleviating downy mildew of cucumber. Int J Plant Anim Environ Sci 2014;4:150–59.
101. Sarhan EA, Abd-Elsyed MH, Ebrahiem AM. Biological control of cucumber powdery mildew (Podosphaera xanthii)(Castagne) under greenhouse conditions. Egypt J Biol Pest Cont 2020;30:1–7. CrossRef
102. Islam MN, Ali MS, Choi SJ, Hyun JW, Baek KH. Biocontrol of citrus canker disease caused by Xanthomonas citri subsp. citri using an endophytic Bacillus thuringiensis. Plant Pathol J 2019;35:486. CrossRef
103. Udayashankar A, Nayaka SC, Reddy M, Srinivas C. Plant growth-promoting rhizobacteria mediate induced systemic resistance in rice against bacterial leaf blight caused by Xanthomonas oryzae pv. oryzae. Biol Cont 2011;59:114–22. CrossRef
104. Al-Rahbi BAA, Al-Sadi AM, Al-Harrasi MMA, Al-Sabahi JN, Al-Mahmooli IH, Blackburn D, et al. Effectiveness of endophytic and rhizospheric bacteria from Moringa spp. in controlling Pythium aphanidermatum damping-off of cabbage. Plants 2023;12:668. CrossRef
105. Xue Y, Zhang Y, Huang K, Wang X, Xing M, Xu Q, et al. A novel biocontrol agent Bacillus velezensis K01 for management of gray mold caused by Botrytis cinerea. AMB Express 2023;13:91. CrossRef
106. Ramírez V, Martínez J, Bustillos-Cristales MR, Catañeda-Antonio D, Munive JA, Baez A. Bacillus cereus MH778713 elicits tomato plant protection against Fusarium oxysporum. J Appl Microbiol 2022;132:470–82. CrossRef
107. Farhaoui A, Adadi A, Tahiri A, El Alami N, Khayi S, Mentag R, et al. Biocontrol potential of plant growth-promoting rhizobacteria (PGPR) against Sclerotiorum rolfsii diseases on sugar beet (Beta vulgaris L.). Physiol Mol Plant Pathol 2022;119:101829. CrossRef
108. Chenniappan C, Narayanasamy M, Daniel G, Ramaraj G, Ponnusamy P, Sekar J, et al. Biocontrol efficiency of native plant growth promoting rhizobacteria against rhizome rot disease of turmeric. Biol Control 2019;129:55–64. CrossRef
109. Perea-Molina PA, Pedraza-Herrera LA, Beauregard PB, Uribe-Vélez D. A biocontrol Bacillus velezensis strain decreases pathogen Burkholderia glumae population and occupies a similar niche in rice plants. Bio Control 2022;176:105067. CrossRef
110. Solano-Alvarez N, Valencia-Hernández JA, Rico-García E, Torres-Pacheco I, Ocampo-Velázquez RV, Escamilla-Silva EM, et al. A novel isolate of Bacillus cereus promotes growth in tomato and inhibits Clavibacter michiganensis infection under greenhouse conditions. Plants 2021;10:506. CrossRef
111. Sultana F, Hossain MM. Assessing the potentials of bacterial antagonists for plant growth promotion, nutrient acquisition, and biological control of southern blight disease in tomato. PLoS One 2022;17:e0267253. CrossRef
112. Hidayah B, Khangura R, Dell B. Biological control potential of Trichoderma species and bacterial antagonists against Sclerotinia sclerotiorum on canola in Western Australia. Int J Agric Biol 2022;27:215–27. CrossRef
113. Kenfaoui J, Lahlali R, Laasli SE, Goura K, Fardi M, Tahiri A, et al. The potency and effectiveness of six essential oils in controlling grapevine trunk diseases in Morocco. J Nat Pest Res 2023;6:100053. CrossRef
114. Delgado A, Toro M, Memenza-Zegarra M, Zúñiga-Dávila D. Control of white rot caused by Sclerotinia sclerotiorum in strawberry using Arbuscular Mycorrhizae and plant-growth-promoting bacteria. Sustainability 2023;15:2901. CrossRef
115. Yadav AN, Kour D, Yadav N. Nano-biofertilizers for agricultural sustainability. J Appl Biol Biotechnol 2023;11:1–4. CrossRef
116. Kour D, Rana KL, Yadav AN, Yadav N, Kumar M, Kumar V, et al. Microbial biofertilizers: biorsources and eco-friendly technologies for agricultural and environmental sustainability. Biocatal Agric Biotechnol 2020;23:101487; CrossRef
117. Shukla K, Negi R, Kaur T, Devi R, Kour D, Yadav AN. First report on rhizospheric silicate mineral weathering bacteria from indian himalayas and their roles for plant growth promotion of tomato (Solanum lycopersium L.). Nat Acad Sci Lett 2023;46:435–38; CrossRef
118. Negi R, Kaur T, Devi R, Kour D, Sheikh I, Tyagi V, et al. First report on Rahnella sp. strain EU-A3SNfb, a plant growth promoting endophytic bacterium from wild wheat relative Aegilops kotschyi. Nat Acad Sci Lett 2022;45:393–96; CrossRef
119. Elnahal ASM, El-Saadony MT, Saad AM, Desoky E-SM, El-Tahan AM, Rady MM, et al. The use of microbial inoculants for biological control, plant growth promotion, and sustainable agriculture: a review. Eur J Plant Pathol 2022;162:759–92; CrossRef
120. Tomar P, Thakur N, Jhamta S, Chowdhury S, Kapoor M, Singh S, et al. Bacterial biopesticides: biodiversity, role in pest management and beneficial impact on agricultural and environmental sustainability. Heliyon 2024;10:e31550; CrossRef
121. Thakur N, Tomar P, Kaur J, Kaur S, Sharma A, Jhamta S, et al. Eco-friendly management of Spodoptera litura (Lepidoptera: Noctuidae) in tomato under polyhouse and field conditions using Heterorhabditis bacteriophora Poinar, their associated bacteria (Photorhabdus luminescens), and Bacillus thuringiensis var. kurstaki. Egypt J Biol Pest Control 2023;33:7; CrossRef
122. Fitriana Y, Tampubolon DAT, Suharjo R, Lestari P, Swibawa IG. Lysinabacillus fusiformis and Paenibacillus alvei obtained from the internal of nasutitermes termites revealed their ability as antagonist of plant pathogenic fungi. Plant Pathol J 2022;38:449. CrossRef
123. Al-Daghari DSS, Al-Sadi AM, Al-Mahmooli IH, Janke R, Velazhahan R. Biological control efficacy of indigenous antagonistic bacteria isolated from the rhizosphere of cabbage grown in biofumigated soil against Pythium aphanidermatum damping-off of cucumber. Agriculture 2023;13:626. CrossRef
124. Negi R, Sharma B, Kumar S, Chaubey KK, Kaur T, Devi R, et al. Plant endophytes: unveiling hidden applications toward agro-environment sustainability. Folia Microbiol 2024;69:181–206; CrossRef
125. Benizri E, Lopez S, Durand A, Kidd PS. Diversity and role of endophytic and rhizosphere microbes associated with hyperaccumulator plants during metal accumulation. In: van der Ent A, Baker AJ, Echevarria G, Simonnot MO, Morel JL (eds.). Agromining: farming for metals. mineral resource reviews, Springer, Cham, Switzerland, pp. 39–279, 2021; CrossRef
126. Kour D, Sharma B, Negi R, Kumar S, Kaur S, Kaur T, et al. Microbial amelioration of heavy metal toxicity in plants for agro-environmental sustainability. Water Air Soil Pollut 2024;235:431; CrossRef
127. Joshi S, Gangola S, Bhandari G, Bhandari NS, Nainwal D, Rani A, et al. Rhizospheric bacteria: the key to sustainable heavy metal detoxification strategies. Front Microbiol 2023;14:1229828. CrossRef
128. Taghavi Ghasemkheili F, Ekelund F, Johansen JL, Pirdashti H, Ghadirnezhad Shiade SR, Fathi A, et al. Ameliorative effects of Trichoderma harzianum and rhizosphere soil microbes on cadmium biosorption of barley (Hordeum vulgare L.) in Cd-polluted soil. J Soil Sci Plant Nutr 2022;22:527–39; CrossRef
129. Shahzad A, Aslam U, Ferdous S, Qin M, Siddique A, Billah M, et al. Combined effect of endophytic Bacillus mycoides and rock phosphate on the amelioration of heavy metal stress in wheat plants. BMC Plant Bio 2024;24:125; CrossRef
130. Backer R, Rokem JS, Ilangumaran G, Lamont J, Praslickova D, Ricci E, et al. Plant growth-promoting rhizobacteria: context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front Plant Sci 2018;9:1473; CrossRef
131. Fadiji AE, Yadav AN, Santoyo G, Babalola OO. Understanding the plant-microbe interactions in environments exposed to abiotic stresses: an overview. Microbiol Res 2023;;271:127368 CrossRef
132. Sylia A, Corrêa A, Cruz C, Yadav A, Nabti E. Plant growth promoting microbes as biofertilizers: promising solution for sustainable agriculture under climate change associated abiotic stresses. Plant Sci Today 2022;8:1–25 CrossRef
133. Kour D, Ahluwalia KK, Ramniwas S, Kumar S, Rustagi S, Singh S, et al. Environment and climate change: influence on biodiversity, present scenario, and future prospect. J Appl Biol Biotechnol 2024;12(2):1–12 CrossRef
134. Kour D, Negi R, Khan SS, Kumar S, Kaur S, Kaur T, et al. Microbes mediated induced systemic response in plants: a review. Plant Stress 2024;11:100334; CrossRef
135. Khoso MA, Wagan S, Alam I, Hussain A, Ali Q, Saha S, et al. Impact of plant growth-promoting rhizobacteria (PGPR) on plant nutrition and root characteristics: current perspective. Plant Stress 2024;11:100341; CrossRef
136. Shahwar D, Mushtaq Z, Mushtaq H, Alqarawi AA, Park Y, Alshahrani TS, et al. Role of microbial inoculants as bio fertilizers for improving crop productivity: a review. Heliyon 2023;9:e16134; CrossRef
137. El-Saadony MT, Saad AM, Soliman SM, Salem HM, Ahmed AI, Mahmood M, et al. Plant growth-promoting microorganisms as biocontrol agents of plant diseases: mechanisms, challenges and future perspectives. Front Plant Sci 2022;13:923880; CrossRef
138. Calvo P, Nelson L, Kloepper JW. Agricultural uses of plant biostimulants. Plant Soil 2014;383:3–41. CrossRef
139. Parray JA, Jan S, Kamili AN, Qadri RA, Egamberdieva D, Ahmad P. Current perspectives on plant growth-promoting rhizobacteria. J Plant Growth Regul 2016;35:877–902. CrossRef
140. Bashan Y, de-Bashan LE, Prabhu S, Hernandez JP. Advances in plant growth-promoting bacterial inoculant technology: formulations and practical perspectives (1998–2013). Plant Soil 2014;378:1–33. CrossRef
141. Schoebitz M, López MD, Roldán A. Bioencapsulation of microbial inoculants for better soil–plant fertilization. A review. Agron Sustain Dev 2013;33:751–65. CrossRef
142. Kumar A, Munder A, Aravind R, Eapen S, Tümmler B, Raaijmakers J. Friend or foe: genetic and functional characterization of plant endophytic Pseudomonas aeruginosa. Environ Microbiol 2013;15:764–79. CrossRef
143. Li GX, Wu XQ, Ye JR. Biosafety and colonization of Burkholderia multivorans WS-FJ9 and its growth-promoting effects on poplars. Appl Microbiol Biotechnol 2013;97:10489–98. CrossRef
144. Berg G, Eberl L, Hartmann A. The rhizosphere as a reservoir for opportunistic human pathogenic bacteria. Environ Microbiol 2005;7:1673–85. CrossRef
145. Allerberger F, Sessitsch A. Incidence and microbiology of salad-borne disease. CABI Rev 2009;4:1–13. CrossRef
146. Compant S, Clément C, Sessitsch A. Plant growth-promoting bacteria in the rhizo-and endosphere of plants: their role, colonization, mechanisms involved and prospects for utilization. Soil Biol Biochem 2010;42:669–78. CrossRef
147. Egamberdieva D, Kamilova F, Validov S, Gafurova L, Kucharova Z, Lugtenberg B. High incidence of plant growth-stimulating bacteria associated with the rhizosphere of wheat grown on salinated soil in Uzbekistan. Environ Microbiol 2008;10:1–9. CrossRef
148. Abhilash P, Dubey RK, Tripathi V, Gupta VK, Singh HB. Plant growth-promoting microorganisms for environmental sustainability. Trends Biotechnol 2016;34:847–50. CrossRef