
1. INTRODUCTION
Textile industries are one among the main cause of water pollution since they require large quantities of water for dyeing processes [1]. It is roughly predicted as 280,000 tons of textile dyes are discharged into different water columns per year as industrial effluent [2]. Azo dyes constitute nearly 70% of overall dyestuffs used for the textile applications which makes it the highest and most commonly used synthetic colorant discharged into the ecosystem [3]. Azo dyes can absorb maximum light from natural visible spectrum due to its chemical structure characterized by one or more azo bonds (–N=N–) [4] and therefore reduces photosynthetic activity, dissolved oxygen concentration, water quality, etc., which causes toxic effects on aquatic flora and fauna and creates severe environmental damages [5].
Azo dyes are extensively utilized by textile industries followed by paper, food, leather, cosmetics, and pharmaceuticals [6]. Untreated and improper treatment on textile dye effluents releases huge amount of liquids containing heavy azo dyes and their toxic metabolites into the aquatic environment which also reveal adverse effects on total biological oxygen demand, chemical oxygen demand, organic carbon, etc. [5]. Recent studies have found that some textile dyes and their toxic metabolites have serious carcinogenic and mutagenic effects on all living systems [7]. Moreover, many reports have evidenced that almost all the textile dyes have toxic effects on the germination rate and biomass production of plant species which are providing a natural habitat for wildlife, preventing soil erosion and improving soil fertility [8]. Based on the above facts, treatment of azo dyes containing effluents and their metabolites is absolutely important before they are discharged into the environment.
Azo dyes are low biodegradable compounds and are challengeable during the wastewater treatment. Such compounds are commonly treated with physicochemical procedures of adsorption, coagulation, and oxidation using ozone. These conventional methods are expensive and produce large amounts of sludge which fails to remove all toxicants and may create secondary pollution [9]. Hence, there is a need on the strategic improvement of a potential, economical, and eco-friendly procedure for the treatment of the textile dyes. In this study, an azo dye, Reactive Blue 194 (RB 194) which has the IUPAC name of pentasodium;4-amino-3-[[5-[4-chloro-6-[3-(2-sulfooxy ethyl sulfonyl) phenyl] imino-1H-1,3,5-triazin-2-yl]-2-sulfonatophenyl] diazenyl] 5-oxido-6-[(4-sulfonatophenyl) diazenyl] naphthalene-2,7-disulfonate, molecular formula, and weight of C33H21ClN9Na5O19S6 and 1190.4 g/mol was studied for its biodegradation activities using a potential dye degrading estuarine bacterium isolated from the Vellar estuary, South India. Further, the potential strain was evaluated for the azoreductase and laccase enzyme production, use of cheaper nutritional substrates for the enhanced activities and ecotoxicity of the dye treated broth.
2. MATERIALS AND METHODS
2.1. Sampling and Isolation of Estuarine Bacteria
Sediment samples were collected from Vellar estuary, Porto Novo, Cuddalore district, Tamil Nadu, India. Samplings were done with the help of sterile spatula and were transferred into pre-sterilized 100 ml bottle containers which were maintained at 4°C in an ice box during transportation and the collected samples were processed immediately in the lab for the isolation of estuarine bacteria. All the possible precautionary measures were applied to minimize the contamination, while collecting and handling the samples. One gram of the collected estuarine sediment sample was serially diluted using pre-sterilized 50% aged sea water and the dilutions were individually spread plated on the yeast glucose agar [10] petri plates prepared using 50% aged sea water. The yeast glucose agar has the composition of yeast extract (10 g/L), glucose (20 g/L), and agar (15 g/L) with the final pH (at 25°C) of 7.5 ± 0.2. After 4 days incubation at 37°C, all the obtained individual colony morphologies were axenic cultured using freshly prepared agar plates under the same applied conditions as described above. All the axenic cultures were stored under lyophilized condition.
2.2. Screening and Molecular Identification of Potential RB 194 Degrading Bacteria
All the collected axenic bacteria were tested individually for their biodegradation activities against a textile azo dye, RB 194 using 50% aged sea water prepared yeast glucose broth supplemented with 100 mg/L of RB 194. The yeast glucose broth has the same composition of yeast glucose agar except agar. After 4 days of incubation at 37°C, the cultured broth samples were centrifuged at 10,000 rpm for 10 min and the absorbance of the cell free supernatants was recorded at λ max (590 nm) of the respective dye using ultraviolet–visible (UV-Vis) spectrophotometer and the uninoculated broth containing the RB 194 dye was used as respective blank. The percentage of dye degradation was calculated using the following formula [11]:
The estuarine bacteria showed that the maximum biodegradation activity was chosen for the further studies. This strain was molecular identified using 16S rRNA partial sequence with the Eubac set of primers, 27F (5’-AGAG TTTG ATCM TGGC TCAG-3’) and 1492R (5’-GGTT ACCT TGTT ACGA CTT-3’) and the phylogenetic tree was plotted using neighbor joining method [12] with highest similar nucleotide sequences available in the NCBI database with the help of MEGA 7 software [13].
2.3. Activities of Azoreductase and Laccase Enzyme during the RB 194 Degradation
The potential strain of this study was cultured in a 1000 mL conical flask with the 400 ml working volume of yeast glucose broth prepared in 50% aged sea water at 37°C temperature, pH 7.5 and 150 rpm agitation as the fermentation conditions. The log growth phase of potential bacterium was used as inoculum which was standardized to 0.1 optical density at 620 nm using UV-Vis spectrophotometer. Activities of the two enzymes, namely, azoreductase and laccase were estimated at 6 h regular intervals till the decline growth phase of the potential bacterium. A portion of cultured broth at the regular time intervals was sonicated using Ultrasonicator (Hielscher, USA) under 20 KHz for 45 s at 4°C and the centrifuged supernatant under 10,000 rpm for 10 min was analyzed for the two enzyme activities with the following assay procedures. Simultaneously, bacterial cell growth and the dye degradation rate were also determined at the same regular intervals as mentioned above. The bacterial growth was evaluated by centrifuging the cell biomass at 10,000 rpm for 10 min followed by the dry biomass at 50°C for 30 min using hot air oven and the biodegradation rate was estimated as per the percentage dye degradation formula used in the screening procedure (section 2.2).
2.3.1. Azoreductase assay
The azoreductase activities were carried out at 30°C in 1 ml reaction volume containing 50 mM sodium phosphate buffer (pH 7.0), 50 μM FMN, 150 μM NADH, 25 μM methyl red (MR), and 200 µl of crude enzyme [14]. The addition of NADH prepared in 50 mM sodium phosphate buffer (pH 7.0) initiated the reaction and enzyme activity was measured with UV-Vis spectrophotometer using the decrease of absorbance at 430 nm per min time. The MR at 430 nm has the extinction coefficient of 23,360 M−1 cm−1. A unit of enzyme activity was expressed as the decrease in the concentration of azo dye (μM) per min.
2.3.2. Laccase assay
The laccase enzyme activities were estimated by 2,2’-azinobis-(3-ethyl benzo thiazoline-6-sulfonic acid (ABTS) method as described by Majcherczyk et al. [15]. The reaction volume has the mixture of 2.450 ml citrate phosphate buffer at pH 4.0, 500 μL of 1.5 mM ABTS and 50 μl crude enzyme which was incubated for 5 min. The oxidation of ABTS was determined by the increase in absorbance at 420 nm (ε420 = 36 mM−1 cm−1) and enzyme activity was represented as Unit/ml/min.
2.4. Influence of Different Carbon, Nitrogen Sources, and Cheaper Edible Flours in the Dye Degradation Activities
The optimization process was carried out under the same fermentation conditions as applied in the enzyme studies and the inoculum was also prepared using the same applied conditions. In this study, different carbon sources such as galactose, sucrose, lactose, and glycogen were replaced for glucose in the yeast glucose broth composition, similarly, various nitrogen sources such as peptone, beef extract, malt extract, and ammonium nitrate were replaced instead of yeast extract which were studied for their highest dye degradation activities. Like the commercial microbiological medium sources, influence of different edible flour substrates such as rice, wheat, corn, and ragi flours at 5% were tested individually for their dye degradation activities instead of all the other nutritional sources. The biodegradation activities were predicted as calculated during the screening of potential bacterium (section 2.2)
2.5. Ecotoxicity Study
The ecotoxicity of the dye treated broth with the potential strain was evaluated using the seed germination rate of Fenugreek (Trigonella foenum-graecum). Experiments were performed on filter paper in petri dishes dipped with the either one of the following toxicity testing solutions, namely, cell free supernatant of dye treated broth, the uninoculated broth without the added dye, solution containing the 100 mg/L dye concentration in distilled water and distilled water alone. In each experiment, ten healthy seeds of equal size were put on the filter paper dipped in 4 ml of toxicity testing sample and the experiments were carried out for 72 h in the dark room. The seed germination rate was estimated using the earlier methodology as described by Kurade et al. [16] and Priac et al. [17] and the radicle growth length of germinated seeds was also evaluated using centimeter scale tape.
3. RESULTS AND DISCUSSION
3.1. Isolation, Screening, and Molecular Identification of the Potential Dye Degrading Estuarine Bacteria
Estuaries are the meeting place of sea and rivers which have delicate ecosystem and are homeland for the unique microbial species. Input of nutrients from tributaries convert the estuarine environment into a highly productive site with tide-dynamics which creates a channel of nutrients, organic matter, waste, etc., from the estuary to the sea [18]. Moreover, physicochemical conditions such as temperature, pH, salinity, and dissolved oxygen concentration are highly variable in estuarine ecosystem due to the flow of freshwater from the rivers which is the main reason behind the formation of unique microbial communities [19]. Unfortunately, estuaries are nowadays being used as a transit for the removal of industrial and agricultural wastes; hence, the adaptive microbial strains of promising detoxicants are highly possible [20]. Hence, an attempt was made in this study for the isolation of a potential dye degrading bacteria from the sediment samples of estuarine environment.
The collected sediment samples were serially diluted using 50% aged sea water and spread plated on 50% aged sea water prepared yeast glucose agar plates for the isolation of estuarine bacteria. After 4 days of incubation, 27 morphologically distinct colonies were isolated and pure cultured repeatedly on the same isolation medium conditions which were initially named as PNS 1–22. All these strains were individually screened for the biodegradation of a textile azo dye, RB 194. After the specified incubation conditions, PNS 15 revealed the maximum activity of 62.8% dye degradation [Figure 1] followed by PNS 7 and PNS 20 with 51.7% and 48.5% dye degradation. Further, 15 strains showed poor activities (0–20%) and nine strains evidenced moderate dye degradation activities (21–40%). Based on the appreciable results, PNS 15 strain was chosen for the further molecular identification and detailed dye degradation studies. Further, only one study was carried out earlier regarding the biodegradation activities of RB 194 using a potential bacterium, Enterobacter sp. isolated from mangrove wetlands of Thailand [21].
The potential strain on this study, PNS 15 was molecular identified based on 16S rRNA partial sequencing method. The polymerase chain reaction amplified sequence revealed the total length of 1550 bp and the BLASTn homology search using the NCBI GenBank nucleotide sequence database revealed 100% sequence similarity with Bacillus megaterium. Based on these results, PNS 15 was identified as B. megaterium and the sequence was deposited in the NCBI GenBank which was provided with the accession number, MT941031.1. The evolutionary relationship of B. megaterium PNS 15 was plotted using neighbor joining method with the highest similar sequences of the NCBI nucleotide collection which was illustrated in the Figure 2. Similarly, the same molecular identification procedure was carried out for the RB 13 degrading bacterium, Proteus mirabilis LAG which was collected from municipal dump site soil near Lagos, Nigeria [22].
3.2. Azoreductase and Laccase Activities of the Potential Strain during the RB 194 Degradation
Azoreductase and laccase are dye degradation enzymes which were investigated for their presence in the biodegradation of RB 194 dye using the potential strain of this study. B. megaterium PNS 15 showed appreciable synthesis of both azoreductase and laccase enzyme. Further, both the enzymes revealed its highest production during the 96th h of incubation period with 0.55 U/ml/min and 1.45 U/ml/min of azoreductase and laccase activities where both the bacterial cell biomass as well as biodegradation rates are also at peak [Figure 3]. The decreased activity of these enzymes reduced the biodegradation rate of RB 194 during the decline growth phase of the bacterium. Furthermore, the degradation of RB 194 by the B. megaterium PNS 15 was initiated from the lag growth phase, simultaneously the synthesis of laccase enzyme was initiated.
 | Figure 1: RB 194 degradation activities during the screening test, (a) showing very poor degradation activity of PNS 4, (b) revealing the highest degradation activity of PNS 15, and (c) distilled water for comparison. [Click here to view] |
Moreover, the dye degradation achieved its peak rate during the stationary growth phase in which both the laccase and azoreductase showed their maximum synthesis, these activities proved the role of these enzyme in the biodegradation of this azo dye, RB 194. In view of enzyme synthesis with reference to the bacterial cell biomass, laccase constituted as growth dependent production whereas azoreductase acts as a secondary metabolite. Similar to this observation, azoreductase and laccase enzyme were synthesized at the highest level during the end of stationary growth phase of the bacterium, Aeromonas punctata in the biodegradation of Reactive Black 5 dye [23]. According to Singh et al. [24], azoreductase and laccase enzyme are the primary sources of any microbes for the appreciable degradation of all classified azo dyes.
3.3. Influence of Different Carbon, Nitrogen Sources, and Cheaper Edible Flours in the Dye Degradation Activities
Standardizing nutritional conditions plays a crucial role in the improvement of biodegradation activities against various toxicants including azo dyes [25]. In this study, various carbon and nitrogen sources were investigated for enhanced biodegradation of the RB 194 dye using the B. megaterium PNS 15. Among the carbon substrates, lactose revealed the highest dye degradation activity of 88.9% followed by galactose, glycogen, and sucrose with 77.2%, 76.5%, and 73.4% biodegradation activities, respectively [Figure 4a]. Regarding nitrogen sources, ammonium nitrate showed the maximum of 93.6% biodegradation activity next by peptone (85.6%), beef extract (79.5%), and malt extract (77.5%), respectively [Figure 4b].
 | Figure 2: Evolutionary relationships of B. megaterium PNS 15 against the highest similar nucleotide sequence of the NCBI database which was inferred based on neighbor-joining procedure. [Click here to view] |
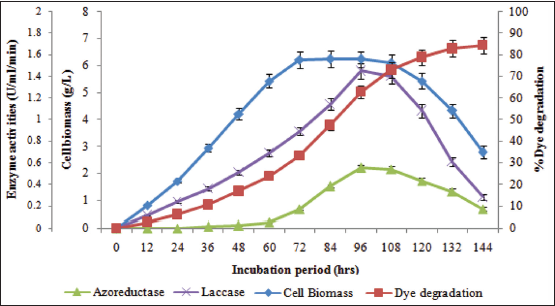 | Figure 3: Azoreductase and laccase enzyme activities of B. megaterium PNS 15 during the RB 194 degradation with reference to cell biomass. [Click here to view] |
Treatment of toxicants using any resources and methodologies has to be economically feasible which make them practically possible in the applied research. In this study, four different cheaper edible flours were studied for their effects on the enhanced degradation activities of the RB 194 dye using the estuarine B. megaterium PNS 15. Interestingly, this strain showed highest biodegradation activity of 92.3% using wheat flour followed by rice (75.6%), corn (72.8%), and ragi (66.8%) flours, respectively [Figure 4c]. Further, the enhanced activity of biodegradation using wheat flour is very close to the biodegradation activity of 93.6% [Figure 5] using the best optimized medium containing the lab biochemical composition of lactose and ammonium nitrate which strongly suggesting the use of wheat flour as the alternative, appreciable, and cheaper nutritional substrate required for the degradation of RB 194 using the potential strain of this study, B. megaterium PNS 15. Similar to this study, wheat bran was used for the biodegradation of RB 172 and Trypan Blue using Providencia rettgeri HSL1 [26] and a microbial consortium [27]. However, many waste substrates have insoluble nature and require pretreatment procedures using toxic chemicals; hence, the present investigation studied on flours for the best treatment options.
3.4. Ecotoxicity Study
The ecotoxicity of the dye degraded broth was studied using the seed germination rate as well as the radical growth length of the T. foenum-graecum. The cell free supernatant of the cultured broth using the cheaper substrate, wheat flour was studied for its ecotoxicity, which revealed 90% seed germination with 2.5 cm radical growth length, whereas the dye solution showed 6.7% seed germination with 0.6 cm radical growth length [Figure 6]. Similarly, the distilled water and uninoculated nutrient broth revealed 100% and 97% seed germination with 2.8 cm and 2.7cm radical growth length, respectively. These observations suggesting the higher non-toxic nature of the RB 194 treated broth using the estuarine bacterium B. megaterium PNS 15 supplemented with the cheaper nutritional sources, wheat flour.
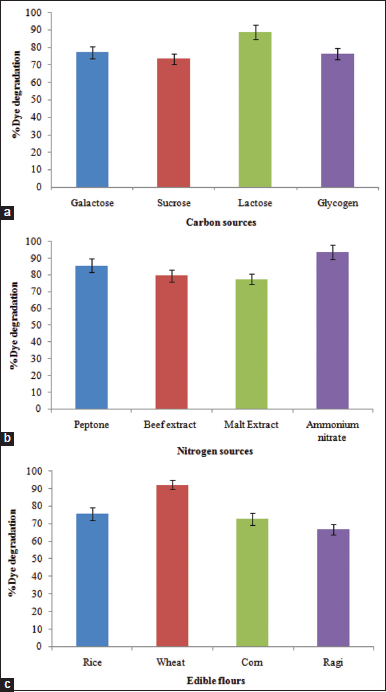 | Figure 4: Influence of different (a) carbon, (b) nitrogen, and (c) cheaper substrates on the RB 194 degradation using B. megaterium PNS 15. [Click here to view] |
 | Figure 5: Appreciable enhanced biodegradation activities of B. megaterium PNS 15 using wheat flour as its nutritional medium substrate. [Click here to view] |
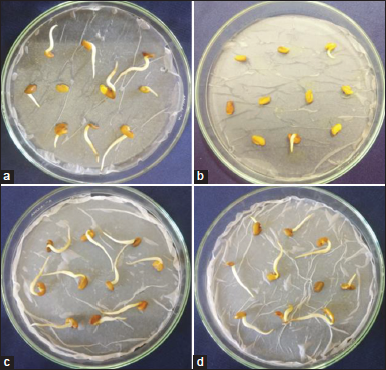 | Figure 6: Ecotoxicity of the samples, (a) cell free supernatant of the treated broth using B. megaterium PNS 15, (b) RB 194 dye solution, (c) distilled water, and (d) uninoculated nutrient broth. [Click here to view] |
In support to this present study, toxicity of the Scarlet R treated broth using two bacterial strains, Proteus vulgaris NCIM-2027 and Micrococcus glutamicus NCIM-2168, was studied with germination rate and radicle growth length of Sorghum vulgare and Phaseolus mungo seeds which revealed no toxicity [28]. Likewise, different azo dyes, namely, Reactive Black 5, Reactive Orange 16, Disperse Red 78, and Direct Red 81 treated broth using a consortium consisted of Providencia rettgeri HSL1 and Pseudomonas sp. SUK1 was investigated for the ecotoxicity using Daphnia magna which also revealed its non-toxic nature [29].
4. CONCLUSION
In this study, a promising RB 194 dye degrading estuarine bacterium, B. megaterium PNS 15 isolated from the sediment samples of an estuarine environment. This potential strain showed an appreciable production of azoreductase and laccase and revealed their significant role in the biodegradation of RB 194. Further, this strain evidenced excellent biodegradation activities of RB 194 using the cheaper substrates, wheat flours as its only nutritional source. Moreover, the dye degraded broth using the B. megaterium PNS 15 with the wheat flour revealed non-toxicity toward the fenugreek seed germination rate which proved its eco-friendly nature. All the above observations strongly suggesting the use of this potential bacterium for the economically feasible and eco-friendly bioremediation of RB 194.
5. CONFLICTS OF INTEREST
The authors declare that they have no conflicts of interest on publication of this article.
6. ACKNOWLEDGMENT
The authors gratefully acknowledge Department of Microbiology, Faculty of Science, Annamalai University, Annamalai Nagar, Chidambaram - 608002, Tamil Nadu, India, for providing lab facilities and supporting our research.
7. AUTHOR CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work. All the authors are eligible to be an author as per the international committee of medical journal editors (ICMJE) requirements/guidelines.
8. FUNDING
There is no funding to report.
9. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
10. PUBLISHER’S NOTE
This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
REFERENCES
1. Banat IM, Nigam P, Singh D, Marchant R. Microbial decolorization of textile-dye containing effluents: A review. Bioresour Technol 1996;58:217-27. CrossRef
2. Jin XC, Liu GQ, Xu ZH, Tao WY. Decolorization of a dye industry effluent by Aspergillus fumigatus XC6. Appl Microbiol Biotechnol 2007;74:239-43. CrossRef
3. Zhao X, Hardin IR. HPLC and spectrophotometric analysis of biodegradation of azo dyes by Pleurotus ostreatus. Dyes Pigments 2007;73:322-5. CrossRef
4. Chang JS, Kuo TS, Chao YP, Ho JY, Lin PJ. Azo dye decolorization with a mutant Escherichia coli strain. Biotechnol Lett 2000;22:807-12. CrossRef
5. Vandevivere PC, Bianchi R, Verstraete W. Treatment and reuse of wastewater from the textile wet-processing industry: Review of emerging technologies. J Chem Technol Biotechnol 1998;72:289-302. CrossRef
6. Telke A, Kalyani D, Jadhav J, Govindwar S. Kinetics and mechanism of reactive red 141 degradation by a bacterial isolate Rhizobium radiobacter MTCC 8161. Acta Chim Slovenica 2008;55:320-9.
7. Lellis B, Fávaro-Polonio CZ, Pamphile JA, Polonio JC. Effects of textile dyes on health and the environment and bioremediation potential of living organisms. Biotechnol Res Innov 2019;3:275-90. CrossRef
8. Kapustka LA, Reporter M. Terrestrial primary producers. In: Calow P, editor. Handbook of Ecotoxicology. Oxford: Blackwell Scientific Publications; 1993. p. 278. CrossRef
9. Sarkar S, Banerjee A, Halder U, Biswas R, Bandopadhyay R. Degradation of synthetic azo dyes of textile industry: A sustainable approach using microbial enzymes. Water Conserv Sci Eng 2017;2:121-31. CrossRef
10. Atlas RM. Handbook of Microbiological Media. 4th ed. Boca Raton, FL: CRC Press; 2010. p. 1932.
11. Alalewi A, Jiang C. Bacterial influence on textile wastewater decolorization. J Environ Protect 2012;3:889-901. CrossRef
12. Saitou N, Nei M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol Biol Evol 1987;4:406-25.
13. Kumar S, Stecher G, Tamura K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol 2016;33:1870-4. CrossRef
14. Qi J, Schlömann M, Tischler D. Biochemical characterization of an azoreductase from Rhodococcus opacus 1CP possessing methyl red degradation ability. J Mol Cataly B 2016;130:9-17. CrossRef
15. Majcherczyk A, Johannes C, Hüttermann A. Oxidation of polycyclic aromatic hydrocarbons (PAH) by laccase of Trametes versicolor. Enzyme Microb Technol 1998;22:335-41. CrossRef
16. Kurade MB, Waghmode TR, Khandare RV, Jeon BH, Govindwar SP. Biodegradation and detoxification of textile dye disperse red 54 by Brevibacillus laterosporus and determination of its metabolic fate. J Biosci Bioeng 2016;121:442-9. CrossRef
17. Priac A, Badot PM, Crini G. Treated wastewater phytotoxicity assessment using Lactuca sativa: Focus on germination and root elongation test parameters. Compt Rend Biol 2017;340:188-94. CrossRef
18. Hoch MP, Kirchman DL. Seasonal and inter-annual variability in bacterial production and biomass in a temperate estuary. Mar Ecol Prog Ser 1993;98:283-95. CrossRef
19. Kimmerer WJ. Effects of freshwater flow on abundance of estuarine organisms: Physical effects or trophic linkages? Mar Ecol Prog Ser 2002;243:39-55. CrossRef
20. Henriques IS, Alves A, Tacao M, Almeida A, Cunha Â, Correia A. Seasonal and spatial variability of free-living bacterial community composition along an estuarine gradient (Ria de Aveiro, Portugal). Estuar Coast Shelf Sci 2006;68:139-48. CrossRef
21. Chantarasiri A. Klebsiella and Enterobacter isolated from mangrove wetland soils in Thailand and their application in biological decolorization of textile reactive dyes. Int J Environ Res Public Health 2020;17:7531. CrossRef
22. Olukanni OD, Osuntoki AA, Kalyani DC, Gbenle GO, Govindwar SP. Decolorization and biodegradation of reactive blue 13 by Proteus mirabilis LAG. J Hazard Mater 2010;184:290-8. CrossRef
23. Roopa KB, Usha MS. Partial purification and assay of azoreductase and laccase in the degradation of reactive black 5 dye from Aeromonas Punctata. Int J Latest Res Eng Technol 2017;3:16-9.
24. Singh RL, Singh PK, Singh RP. Enzymatic decolorization and degradation of azo dyes-a review. Int Biodeter Biodegrad 2015;104:21-31. CrossRef
25. Bardi L, Marzona M. Factors affecting the complete mineralization of azo dyes. In: Biodegradation of Azo Dyes. Berlin, Heidelberg: Springer; 2010. p. 195-210. CrossRef
26. Lade H, Govindwar S, Paul D. Low-cost biodegradation and detoxification of textile azo dye CI reactive blue 172 by Providencia rettgeri strain HSL1. J Chem 2015;894109. Doi.org/10.1155/2015/894109. CrossRef
27. Lade H, Kadam A, Paul D, Govindwar S. A low-cost wheat bran medium for biodegradation of the benzidine-based carcinogenic dye trypan blue using a microbial consortium. Int J Environ Res Public Health 2015;12:3480-505. CrossRef
28. Saratale RG, Saratale GD, Kalyani DC, Chang JS, Govindwar SP. Enhanced decolorization and biodegradation of textile azo dye Scarlet R by using developed microbial consortium-GR. Bioresour Technol 2009;100:2493-500. CrossRef
29. Lade H, Kadam A, Paul D, Govindwar S. Biodegradation and detoxification of textile azo dyes by bacterial consortium under sequential microaerophilic/aerobic processes. EXCLI J 2015;14:158-74.