
1. INTRODUCTION
Heavy metal contamination of soil, particularly Copper (Cu) accumulation, represents a significant environmental concern due to its persistence, toxicity and potential for bioaccumulation [1]. Cu is the 25th most prevalent metal in the crust of the Earth. This metal is frequently found at high levels in textile effluent water, and this poses significant ecological and health risks due to its toxicity [2]. It is considered a vital micronutrient necessary for the growth and development of plants. Although Cu is an essential micronutrient required for the plant and microbial metabolism, excessive Cu present in the soil can be hazardous and extremely poisonous to humans, animals, flora, and microbiota [3]. Industrial activities, including textile processing, contribute substantially to Cu discharge into surrounding environments, leading to long term soil contamination [4].
Conventional approaches, such as biological, chemical, and physical techniques are often employed to remove Cu from wastewater, but are often inadequate to decrease the toxicity in soil [5]. Consequently, biological approaches, particularly microbial bioremediation, have gained attention as sustainable and environmentally compatible alternatives. Among them, using biosurfactants to remediate heavy metals is an eco-friendly and practical way to get clean soil [6]. Numerous microorganisms generate these biosurfactants that have surface-active properties [7]. These amphiphilic biomolecules, produced by a variety of bacteria such as Bacillus, Pseudomonas [8], and Gordonia [9], reduce surface and interfacial tension, form micelles, increase metal mobility, and aid in biodegradation processes [8-10]. These biosurfactants cause hydrocarbons to pseudo-solubilize by forming micelles [9]. Toxic contaminants, including Cu, lead (Pb), chromium (Cr), cadmium, and others, are broken down by these biologically generated surfactants. The process of metal remediation and the creation of the biosurfactant-metal complex are extensively reported by microorganisms, and also by similar complexes [9]. These bioactive molecules are primarily characterized by low toxicity, ecological acceptability, and high biodegradability, making them viable substitutes for chemically manufactured surfactants and active in metal reduction/removal. The biosurfactant [10] extracted from the Bacillus cereus strain NWUAB01 formed a complex with metal and reduced cadmium (Cd), Pb, and Cr [11] from polluted sites contaminated with oil.
The non-ribosomal peptide synthase (NRPS) gene involved in the synthesis of a lipopeptide biosurfactant “surfactin,” showcase the presence of thioesterase domain protein (SrfTE) and the Cu-exporting P-type ATPase (CopA). CopA is reported responsible for Cu metal bioremediation [12]. The SrfTE domain plays a crucial role in biosurfactant activity by facilitating chain termination, macrocyclization, and the release of surfactin. SrfTE domains are highly conserved and often used as representative markers in partial cluster analyses [13,14]. Surfactin enhances metal desorption, solubilization, and soil mobility [15,16]. The CopA performs vital functions in the translocation of Cu [Cu(I)] ions across the cell either for the biosynthesis of Cu proteins or detoxification of the Cu ions from the surroundings [17]. Both proteins have distinct pathways within bacterial cells [18]. Bacteria possessing these two genes can be used for bioremediation of effluent sites by removing metal contamination caused by various metals, such as Cu, zinc, and cadmium [19]. However, studies integrating the genomic identification of these determinants with experimental validation of soil-level Cu reduction are limited.
Therefore, the present study is aimed (i) to characterize the biosurfactants produced from Bacillus paramycoides strain NT001 isolated from Cu contaminated soil, (ii) evaluate the Cu remediation in soil, and (iii) investigate the presence of the gene NRPS associated SrfTE domain, and CopA ATPase, genes to establish the genetic potential underlying biosurfactant production and Cu tolerance.
2. MATERIALS AND METHODS
2.1. Chemicals
Chemicals, reagents, and solvents were purchased from Himedia Laboratories Private Limited (Mumbai, Maharashtra, India) and Sisco Research Laboratories Pvt Ltd. (SRL, Mumbai, Maharashtra, India). Standard Surfactin (90% purity) and Dimethyl sulfoxide d6 were procured from Sigma-Aldrich (St. Louis, MO, USA). Crude oil was purchased from the fuel pump station. The chelating agent diethylene triamine penta acetic acid (DTPA) (Sigma) was utilized exactly as supplied. The study’s metal-containing soils were procured from the contaminated location in Sanganer, which were air-dried for a month before being sieved through a 2 mm screen. Typical studies were carried out with a batch solution volume (V) of 100 mL in 125 mL glass Erlenmeyer flasks.
2.2. Microorganism
The novel bacterial strain considered for this study was isolated from the textile industry-contaminated soil from Sanganer (Latitude 26.7° N and Longitude 75.7°E), Jaipur, Rajasthan [20].
2.3. Culture medium
The bacterial isolate was isolated [20], using serially diluted effluent contaminated soil (1 g) using modified minimal salt media [MSM] supplemented with 1% diesel as a carbon source. The media was composed of 3.6 g/L of Na2HPO4; 1 g/L of (NH4)2SO4; 1 g/L of KH2PO4; 1 g/L of MgSO4·7H2O; 0.01 g/L of Fe(NH4) citrate; 0.1 g/L of CaCl2·2H2O; 0.1 g/L of yeast extract; and 20 g/L of glucose. Along with 0.1% of the trace element stock solution, it was added. The trace element solutions consisted of g/L ZnSO4·7H2O; 3 μg/L MnCl2·4H2O; 1 μg/L CoCl2. 6H2O; 2 μg/L NiCl2·6H2O; 3 μg/L Na2MoO4·2H2O; 30 μg/H3BO3; and 1 μg/L CuCl2·2H2O. The pH of the medium was adjusted to 6.8 and it was incubated at 37°C and 120 rpm in an incubator for 7 days. The microorganism was streaked over nutrient agar media to isolate the pure cultures, which were then cultured for 48 h at 37°C [21]. Later, the cultures were maintained on nutrient agar. It was identified as B. paramycoides strain NT001, with National Centre for Biotechnology Information (NCBI) Accession number MK613839. This identification was based on the similarity with B. paramycoides strain MCCC 1A04098 (Accession number NR_157734.1), which served as the reference sequence for performing basic local alignment search tool (BLAST) and phylogenetic analysis using the consensus sequence generated from 16S ribosomal ribonucleic sequencing [Figure S1] [19,22].
2.4. Bacterial growth assessment and biomaterial biosurfactant production by B. paramycoides strain NT001
A loopful from a fresh culture of B. paramycoides NT001 [Figure S2a] grown on nutrient agar was inoculated into nutrient broth and incubated at 37°C and 150 rpm for 3 days. The bacterial growth analysis was performed by the gravimetric method by measuring the biomass of the bacterial culture. The cultures were retrieved as pellets after every 24 h by centrifugation for 15 min. at 4°C and 10,000 rpm. This process was continued for 3 days. The pellets were further dried in the oven and weighed for assessment of biomass. The supernatant underwent acidification to pH 2 using 6 N HCl, followed by overnight chilling treatment at 4°C for precipitation of the crude biosurfactant [Figure S2b]. Chloroform and methanol were used in the ratio 65:15 (v/v) and shaken vigorously in a separating funnel for liquid-liquid separation [Figure S2c]. Finally, the partially purified biosurfactant was dried at 40°C in a rotary evaporator and later stored at 4°C. The net weight (g/L) was measured and subsequently characterized to comprehend the key features of the biosurfactant [20].
2.5. Physico-chemical characterization of biosurfactant-producing bacteria
2.5.1. Emulsification
The activity of emulsification of B. paramycoides NT001 was verified using diesel. In this test, 2 mL cell-free supernatant was added to the same amount of diesel and vortexed for 1 min, and allowed to stand for 3 days. After every 24 h of the period, the height of the emulsion and the total height were measured. The emulsification index (E24) was computed by multiplying the ratio of the emulsion layer height to the overall height by 100 [20].
2.5.2. Surface tension
The surface tension of the cell-free supernatant of B. paramycoides NT001 grown for 24, 48, and 72 h was measured using a tensiometer (Drop shape analyzer-DSA25, KRUSS) to verify its ability to reduce the surface tension [22].
2.5.3. Critical micelle concentration (CMC) of biosurfactant
The CMC extracted biosurfactant of B. paramycoides NT001 was analyzed by the pendant drop method using a Goniometer (Dataphysics OCA 15EC). To measure the CMC, concentration variables of different dilutions were prepared, and their interfacial tension was noted. The breakpoint of interfacial tension and biosurfactant concentration was used to analyze the CMC of the material [20].
2.5.4. Particle size and zeta potential estimation of biosurfactant
To measure the particle size and electrokinetic potential of the extracted biosurfactant of B. paramycoides NT001, the crude biosurfactant suspension was filtered through a 0.22 µm filter. An Anton Paar Litesizer 500 was used to analyze the size and charge of the extracted biosurfactant [20].
2.6. Characterization to understand the chemical nature and type of biosurfactant
2.6.1. Fourier transmission infrared spectroscopy (FTIR)
The chemical composition of the recovered biosurfactant of B. paramycoides NT001 through KBr pelleting was evaluated using FTIR spectroscopy. An FTIR spectroscope (Bruker Optics, Ettlinger, Germany, Model Alpha-T with Eco ATR) was used to analyze a fine dry powder made from 1 mg of the dried crude biosurfactant and 100 mg of KBr powder between 4,000 and 400 cm−1 [20].
2.6.2. Triple quadrupole liquid chromatography-tandem mass spectrometry (LC-MS/MS)
The LC-MS/MS analysis was performed using an Agilent 6470 triple quadrupole system. Instrument parameters were set at 300°C gas temperature, 10 L/min sheath gas flow, 3,500 V capillary voltage, 30 psi nebulizer pressure, and a mass range of 100–1,200 m/z in positive ion mode. Eluent A (water) and Eluent B (methanol) were used in a gradient program as described previously [21]. A primary stock solution of surfactin standard (1 mg/mL) was prepared and serially diluted to obtain a 7-point calibration curve within the linear range of 5–500 ng/mL. All calibration standards were freshly prepared in methanol. The reported concentrations represent back-calculated values after applying dilution factors. Before LC-MS/MS analysis, the biosurfactant sample extracted from strain NT001 was filtered and diluted appropriately in methanol and sonicated for 30 min to ensure complete solubilization. It was to ensure that measured biosurfactant concentrations fell within this calibration range [21].
2.7. Bioremediation of Cu present in textile effluent-contaminated soil
2.7.1. Methodology
These biosurfactants form micelles and cause hydrocarbons to pseudo-solubilize [9]. These naturally occurring surfactants facilitate the degradation of harmful contaminants found in wastewater, including Pb, Cu, Cr, Cd, and many more. A 24-h culture of B. paramycoides NT001 (optical density 0.8–1.0 and a colony-forming unit of 108–1010) was added to the polluted soil (5 kg each for different sets), appropriately mixed, and allowed to dry in the sun for 10 days. This procedure was carried out twice. The same soil was further treated using a crude biosurfactant derived from the bacterial isolate NT001. Normal garden soil was also used in this experiment as a positive control (pH 6.9–7.5 and organic matter 3–6%).
2.7.2. Elemental profiling and quantification of Cu
The dehydrated samples were also analyzed using energy-dispersive X-ray spectroscopy (EDX), AMTEX, USA, and the EDX APEXTM V1.3.1 algorithm at 20 kV. The samples were placed on carbon tapes and given a gold coating for 30 s before being studied by a field emission scanning electron microscope, JEOL manufacturer JSM-7610FPlus, Japan, working at 5 kV. Using EDX, the elemental composition of the soil was ascertained.
Quantification of Cu in contaminated and treated soil was initiated by pretreatment as per the protocol of the DTPA extraction method [24]. A total of 5 kg of soil from both sets was separately combined with DTPA, agitated for 2 h, and filtered. To estimate the amount of Cu present in the soil samples, the filtered soil samples from both sets were digested and diluted for further use during quantification. They were measured using a Photon K lamp and a Flame Atomic Absorption Spectrometer (AAS-ECIL manufacturing company). Before the commencement of the experiment, all calibrations using standard solutions were completed [20]. The initial concentration of Cu in the contaminated soil, as well as the remediated soil sample after treatment with bacteria NT001 and its biosurfactant, was calculated after calculating the Cu mass balance [13,14]. The mass of the soil multiplied by the original concentration of the Cu yielded the initial mass of Cu. Using the final concentration, the remaining mass of Cu following treatment was further computed. The amount of Cu liberated (removed) into the DTPA extracts was indicated by the difference between the initial and residual mass. The original mass is equal to the sum of the released and remaining mass, according to the mass balance. Finally, the percentage of Cu reduction relative to the starting mass of Cu in the contaminated soil was computed [25].
2.7.3. Bioinformatic study of NRPS and CopA in B. paramycoides NT001
Phylogenetic analysis was performed to determine the evolutionary placement of the strain NT001 within the genus and assess the relatedness to reference B. cereus complex. To elucidate the phylogenetic relatedness and homology of the thioesterase domain (SrfTE) within the NRPS cluster of closely related Bacillus species, an in silico analysis was conducted for the biosurfactant-producing bacterial strain B. paramycoides NT001 [15,26]. The isolate studied for the present investigation, B. paramycoides NT001, lacks the availability of an assembled or annotated whole genome. Therefore, full operon-level analysis (srfA-A, srfA-B, srfA-C, srfA-D) could not be performed. The SrfTE domain was chosen for this study as it is essential for producing mature surfactin [26] and also helps predict biosurfactant-mediated remediation potential [27].
Search phrases and the Boolean operator “NRPS AND bacillus” were used to extract protein sequences from the NCBI database. Multiple sequence alignment and UniProt were used to gather amino acid sequences using CLUSTAL-W Omega software [28]. Representative NRPS protein sequences were retrieved from the NCBI database based on the highest similarity to the NT001 NRPS cluster and functional annotation. Redundant and partial sequences were excluded to ensure accurate phylogenetic comparison. The analysis involved 11 amino acid sequences for the thioesterase domain, respectively.
MEGA 11 software version 11.0.13 was used to perform multiple sequence alignment for these proteins [29]. An evolutionary cladogram [Figure 1a] for the thioesterase domain of NRPS and the CopA was created. The method for alignment was performed using CLUSTAL-W Omega software [28]. Furthermore, a phylogenetic analysis was conducted where rectangular cladograms were inferred using the Maximum Likelihood method for thioesterase [28] based on the JTT matrix-based model [30]. The frequency of clustered taxa is shown in red on branches.
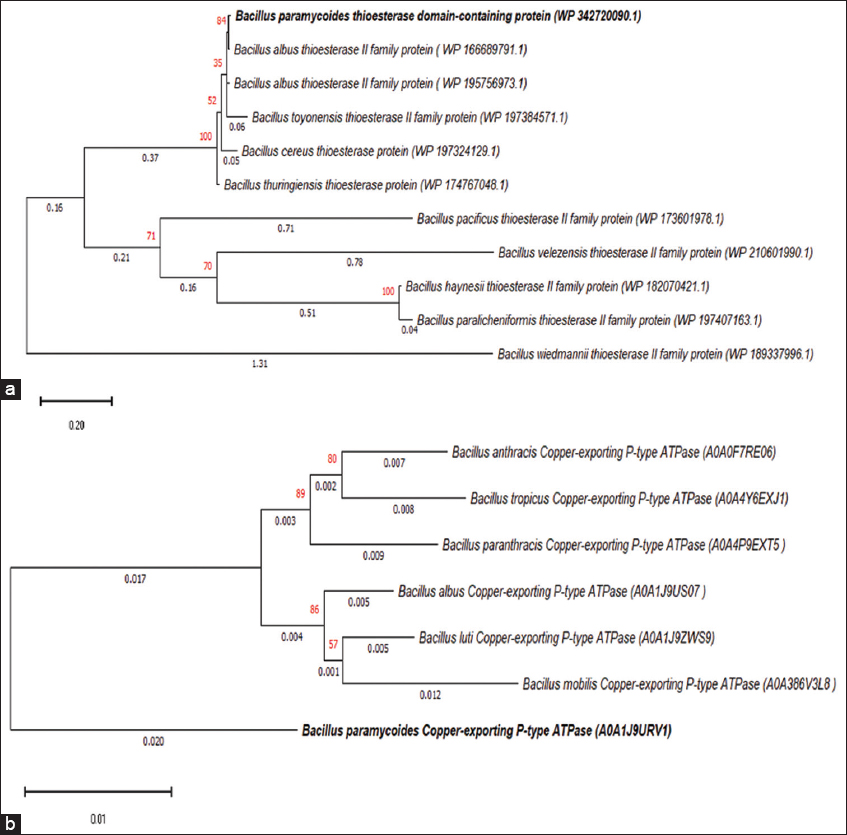 | Figure 1: Phylogenetic tree constructed using MEGA 11 software for amino acid sequences. The tree was generated by the Neighbor-Joining method with 1000 bootstrap replicates to show evolutionary relationships among homologous proteins. The sequences corresponded to (a) thioesterase domain present in non ribosomal peptide synthetase -(Accession No. WP 342720090.1) gene cluster, responsible for surfactin biosynthesis, and (b) Copper-exporting P-type ATPase in Bacillus paramycoides (A0A1J9URV1) and similar Bacillus species. [Click here to view] |
Similarly, in silico studies for the detection of CopA for the same isolate B. paramycoides were performed to assess the potential of the bacteria to bioremediate Cu. Protein sequences were extracted from the NCBI database by employing search terms and using the Boolean operator “Copper P-type ATPase,” “Heavy metal P-type ATPase,” AND bacillus.” The method for alignment and the phylogenetic analysis for CopA was performed in the same manner as mentioned for NRPS. CopA homologs were selected based on protein-BLAST similarity search against NCBI databases. Only full length functionally annotated P type ATPase Cu efflux proteins from representative Bacillus species were included. After removal of redundant entries, the major impressions study involved seven amino acid sequences for the CopA domain, for 7 sequences for phylogenetic analysis.
3. RESULTS
3.1. Characterization of biosurfactant-producing B. paramycoides NT001
The preserved bacterial strain, B. paramycoides strain NT001, revived on modified MSM media, showed similarities as described in our earlier report [20]. When cultures were streaked on nutrient agar medium, their morphological presence was noted after 24 h of incubation. They appeared as white colored colonies, irregular, with an undulated margin, raised elevation, moist texture, shiny appearance, with translucent optical property [Figure S1a]. Using an acid precipitation method, biosurfactant is extracted from the isolated strain NT001 [Figure S1 b and c].
The temporal dynamics show the changes in surface tension, emulsification, biomass accumulation, and yield of biosurfactant from the supernatant over time. The maximum biomass of 2.62 g/L was observed after 72 h. The rise in growth of B. paramycoides strain NT001, and the biosurfactant yield, suggested a relationship between bacterial biomass growth and biosurfactant production. With the increase in time, the surface tension decreases after 72 h. The surface tension of the extracted biosurfactant reduced from 72 mN/m to 45.37 mN/m. It also formed emulsions with diesel. The emulsification index [E24] with diesel was checked and found to range from 33.33% to 42.02%, as shown in Figure S3. All these tests confirmed that our novel isolate NT001 is a potential biosurfactant producer.
3.2. Physicochemical characteristics of the extracted biosurfactant
3.2.1. CMC of biosurfactant
The surface tension measurements at different concentrations resulted in the lowest concentration required to form the micelle, the CMC of the biosurfactant. The surface tension of a novel biosurfactant extracted from the B. paramycoides strain NT001 decreased from 72 to 47.88 mN/m at a concentration of 4 mg/mL, as shown in Figure 2a, and remained constant even after increasing the concentration to 10 mg/mL. Thus, the CMC of the biosurfactant extracted was 6 mg/mL. It is understood that from the point of CMC, biosurfactants initiate self-aggregation. For bioremediation, surfactants with lower CMC values are recommended since solubilization only works at surfactant concentrations higher than the CMC [31].
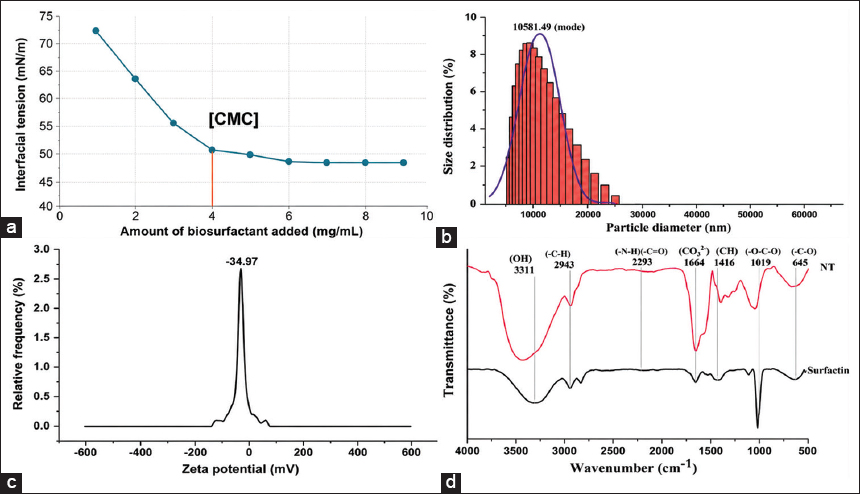 | Figure 2: Physicochemical and functional characterization of the crude biosurfactant produced by Bacillus paramycoides strain NT001 isolated from the Sanganer area in Jaipur. (a) Critical micelle concentration; (b) Dynamic light scattering; (c) Zeta potential; (d) Fourier transmission infrared spectroscopy showed similarity with standard surfactin. [Click here to view] |
3.2.2. Particle size and zeta potential estimation of biosurfactant
The dynamic light scattering technique was used to measure particle size of the biosurfactant, and the hydrodynamic diameter was observed at 1081.40 nm [Figure 2b]. The biosurfactant extracted from B. paramycoides strain NT001 was diluted, ensuring high transmittance (76.3%). Polydispersity index and diffusion coefficient were observed to be 32.7% and 0.5 μm2/s, respectively. Transmittance from the solution was 59.0% while conductivity was 0.173 mS/cm. Overall charges on biosurfactant surface were observed as (−) 34.27 ± 0.5 mV [Figure 2c]. Due to its negatively charged hydrophilic group, the biosurfactant is categorized as an anionic surfactant.
3.3. Functional characterization of the biosurfactant
3.3.1. Fourier transform infrared spectroscopy
A distinct similarity between the functional groups of the biosurfactant extracted from the isolate NT001 and those of the commercial standard surfactin was observed. In [Figure 2d], the peaks correspond to the stretching vibrations of the hydroxyl (OH) group at 3,311 cm−1, aliphatic (C-H) at 2,943 cm−1, and nitrile (C ≡ N) at 2,203 cm−1, respectively. At 1,654 cm−1, a strong absorption band of the CO-N group symbolizes the presence of the stretching vibrations and peptide bonds characteristic of surfactin. The CH group of the bending/stretching of the aliphatic chain was noted at 1,430 cm−1. The ester carbonyl group, or O-C-O of stretching vibrations, was detected at 1,019 cm−1, and the bending vibration of C=C noted at 643 cm−1, which predicts the alkene group [32]. On comparison of the peaks, it was noted that this extracted biosurfactant may be surfactin [21].
3.3.2. Chemical characterization of a biosurfactant by LC-MS/MS
Biosurfactant extracted from B. paramycoides strain NT001 was further identified by LC-MS/MS. The MS scan of the standard surfactant type of biosurfactant “surfactin” was compared with the extracted biosurfactant NT001 according to MS peak [M + H] profiles. The conventional total on chromatogram of multiple reaction monitoring scan of the precursors was observed at 1036.70 (9.79 min) in [Figure 3a]. When collision-induced dissociation occurred, the standard surfactin fragmented and produced a daughter ion with an m/z of 685 [Figure 3d]. It was found in both the extracted biosurfactant from B. paramycoides strain NT001 and standard surfactin surfactant [Figure 3b and c]. A linear curve with a correlation coefficient of 0.994 was seen between 5 and 500 ng/mL [21]. The quantity of surfactin-type lipopeptide biosurfactant extracted from B. paramycoides strain NT001 was quantified and found to be 162.37 ng/mL.
 | Figure 3: Identification of surfactin in the partially purified biosurfactant extracted from Bacillus paramycoides strain NT001 using a triple quadrupole liquid chromatography-mass spectrometer. (a) Total ion chromatogram of standard surfactin. Retention time of (b) standard surfactin and (c) extracted biosurfactant. (d) daughter ion of strain NT001 with a similar daughter ion mass of 685.3 as in standard surfactin. (e) 1H Nuclear magnetic resonance spectra with chemical shift similar to standard surfactin. [Click here to view] |
3.3.3. NMR of the biosurfactant
The 1H NMR spectra [Figure 3e] are consistent with surfactin [23,33]. Prominent signals are observed in the aliphatic region, especially between δ 0.7 and 1.5 ppm, corresponding to methyl and methylene protons, probably within the lipid tail. Near the peptide ring structure, δ 3.5 and 4.0 ppm, characteristic protons were noted. Additional signals near the carbonyl group, δ 2.0 and 2.5, may be due to methylene and methine groups [21].
3.4. Bioremediation of textile effluent-contaminated soil with biosurfactant for the reduction of Cu from contaminated soil
3.4.1. Functional genomic analysis of metal resistance and biosurfactant synthesis genes
The phylogenetic cladogram demonstrated that the strain NT001 clusters with members of B. cereus group, supporting taxonomic placement within this complex. This clustering aids in providing an evolutionary context for the presence of primary P-type ATPase (CopA) responsible for active efflux from the cytoplasm in Gram-positive bacteria. Thus is widely recognized as the central determinant of the bacterial Cu tolerance [34]. Similarly, the surfactin biosurfactant-producing genes (NRPS gene systems), are widely conserved among members of this clade. In our in silico analysis, it was revealed by the evolutionary relatedness of both the thioesterase domain of NRPS and the CopA in B. paramycoides using arbitrary sequences extracted from NCBI and UniProt databases. During this analysis, multiple sequence alignments revealed identity between the query protein sequence and that is the SrfTE present in the NRPS of B. paramycoides (Accession No. WP 342720090.1) and other subject sequences [Figure S3 and 4]. From Figure 1a, the cladogram shows that the SrfTE domain of B. paramycoides (Accession No. WP 342720090.1) has the least evolutionary distance (0.008) with Bacillus albus (Accession No. WP 166689791.1), as they share the same clade. In contrast, Bacillus wiedmannii has the maximum divergence (branch length 1.311) and is distantly placed from B. paramycoides. In Figure 1b, the cladograms showed a similarity among the species. B. albus and Bacillus luti showed the least evolutionary distance (0.005) from the root sequence B. paramycoides. In contrast, Bacillus mobilis shows the maximum branch length of 0.012.
3.4.2. EDX and AAS analysis of soil
Analysis using EDX of textile effluent-contaminated soil, both before and after treatment with B. paramycoides strain NT001 and its surfactin biosurfactant, showed a differential pattern of distribution and concentration of the elemental composition, including carbon, oxygen, and Cu. It indicates changes in the soil’s chemical composition [Figure 4a-c]. Using EDX analysis, it was found that there was an increase in oxygen from 27% to 35% in weight percentage and 28–37.6% in atomic elemental percentage [Figure 4d]. As seen in Table 1, the decrease in carbon content from 45% to 31% in weight % and the similar decrease in atomic percentage from 63% to 45% are indicative of the degradation/breakdown of the carbon-rich compounds, leading to a decline in carbon content. According to a Cu mass balance measurement, the 5 kg soil sample had 3.935 mg of Cu before the treatment. After treatment, 2.035 mg of Cu was released into the DTPA extracts, while 1.900 mg of Cu remained in the soil samples. Thus, a 51.7% decrease in Cu content was noted in the soil after bioremediation. One-way analysis of variance and mean comparison tests regarding the Cu content across the samples showed that the treatment had a significant effect on the Cu content in the soil (F = 94.57, P < 0.001). Contaminated soil shows the highest Cu concentration and is statistically different from normal soil and treated soil. Subsequent comparisons revealed that the treated soils grouped with the normal soils (both labeled “b”), confirming the effective Cu reduction by NT001, and on the contrary the degraded soils formed a separate group (“a”), indicating significantly higher Cu concentrations. These results demonstrate that NT001 treatment decreased Cu accumulation effectively [Figure 4c and Table S2].
Table 1: Elemental composition of textile effluent-contaminated soil obtained from EDX data.
| Soil | Element | Weight % | Atomic % of elements |
|---|---|---|---|
| Soil | C K | 3.6 | 6.3 |
| O K | 45.7 | 60.3 | |
| Cu K | 0.7 | 0.2 | |
| Contaminated | C K | 45 | 63.3 |
| O K | 27 | 28 | |
| Cu K | 11 | 3.1 | |
| Treated | C K | 31.4 | 45 |
| O K | 35 | 37.6 | |
| Cu k | 8.8 | 11.6 |
EDX: Energy-dispersive X-ray spectroscopy
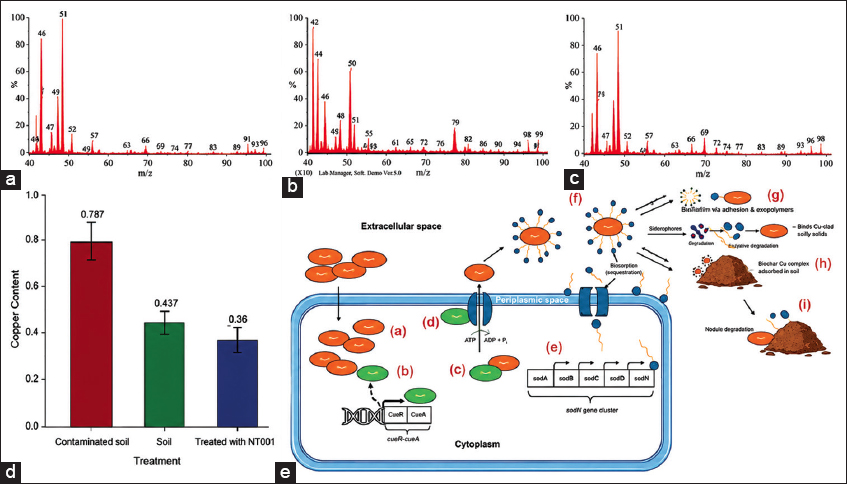 | Figure 4: Bacillus paramycoides NT001 and its biosurfactant-assisted bioremediation of copper in contaminated soil, based on investigations using energy dispersive X-ray spectroscopy of (a) Control, (b) Contaminated, (c) Treated, (d) Quantification of copper using atomic absorption spectrometer. (e) The Cop operon and NRPS cluster are schematic and conceptual models rather than experimentally demonstrated in this strain. The copper metal efflux mechanism in the bacterial cell involves the (a) Cu+2 ions uptake from the extracellular environment, (b) binding of CsoR to Cu+2, inducing CopZ expression, (c) CopZ binds to Cu+ ions and attaches to (d) CopA. (e) Surfactin, produced by (f) NRPS gene cluster, solubilize Cu+2 ions, facilitating (g) bioremediation through disintegration, (h) biodegradation, or (i) biosorption mechanisms [Click here to view] |
4. DISCUSSION
The biosurfactant produced by strain NT001 exhibited surfactant capabilities, including foaming, detergency, and emulsification, and exhibited anionic nature [35]. It was characterized as lipopeptide using FTIR, LC-MS/MS, and 1H NMR [21,33] and found to be similar to “Surfactin” [36-38]. Previous studies using Bacillus species that produced surfactin or pure biosurfactants in lab settings have found Cu removal efficiencies ranging from 5% to 40% removal, depending on matrix complexity and experimental design [36-40]. In contrast, the Cu reduction found in our present investigation using strain NT001 and its biosurfactant treatments falls beyond this range, suggesting better remediation under soil circumstances. Due to its “anionic” nature, this surfactin could interact with positively charged metal matrices, including Cu2+ though chelation and electrostatic attraction. This facilitates the reduction of Cu present in contaminated soil by increase in could interact and bioavailability of Cu present in soil [35]. This increases carbon intake during growth and metabolism, as well as the biodegradation of carbon compounds during decomposition. Thereby implies that adding bacteria that produce biosurfactants to a polluted soil will result in an improvement in the biodegradation. These bacteria also have enzymes that metabolize and produce carbon dioxide and water [38]. This may be related to the oxidation reactions occurring during the active biodegradation process at the site of treatment by the microorganism [39]. In this present investigation, the efflux of metal by exporters and bio-removal of metals by biosurfactant can be hypothetically proposed as methods of removal. A genomic approach involving CopA and SrfTE, present in the NRPS, can be responsible for Cu metal bioremediation [10,41-47]. Genomic analysis further revealed the presence of CopA gene, which is known to provide intracellular Cu tolerance by exporting excess Cu2+ from the cytoplasm.
The role of CopA in remediation deals with the export of excess Cu2+ from the cytoplasm to the periplasm, enabling bacterial survival in metal-rich soil [12,16]. The basic process of translocating Cu ions outside the cells is depicted in Figure 4e [42-47]. Thus, we believe that the combined evidence and experimental reduction data support the potential strain NT001 as a candidate for Cu bioremediation, while recognizing that detailed mechanistic studies remain a scope for further investigation.
5. CONCLUSION
This is the first report of the novel bacteria, B. paramycoides strain NT001, isolated from the textile effluent-contaminated site, that has been extensively characterised to produce biosurfactant “surfactin.” This surfactin aided in the formation of a micelle by self-aggregation. The low particle size of biosurfactants and their anionic charges assisted in the bioremediation of Cu from contaminated soil. The processes enabling Cu detoxification in this strain NT001 were clarified by the involvement of thioesterase and CopA in the molecular pathways. In the future, gene knockout studies or transcriptional analysis (reverse transcription polymerase chain reaction) can offer more immediate functional validation. Thus, the dual presence of both the bacteria and the biosurfactant portrayed the facilitated bioremediation of the Cu from the contaminated soil. A combinatorial use of the surfactin-producing organism and surfactin can pave the way to lower the concentration of pollutants that are hazardous to the ecosystem.
6. ACKNOWLEDGMENT
The authors acknowledge the instrumental support provided by the Sophisticated Analytical Instrumentation Facility and the Central Analytical Centre at Manipal University, Jaipur. MD expresses gratitude for the support received from the Manipal Research Board (MRB seed grant, DoR MRB|2023|SG-03) and is also thankful for the grant awarded to the Department of Biosciences at Manipal University, Jaipur, funded by the Department of Science and Technology (DST-FIST project No. DST/2022/1012), Government of India.
7. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work. All the authors are eligible to be author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
8. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
9. ETHICS APPROVAL
This study does not involve experiments on animals or human subjects.
10. DATA AVAILABILITY STATEMENT
The data that support the findings of this study are available from the corresponding author on reasonable request.
11. PUBLISHER’S NOTE
All claims expressed in this article are solely those of the authors and do not necessarily represent those of the publisher, the editors and the reviewers. This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
12. USE OF ARTIFICIAL INTELLIGENCE (AI)-ASSISTED TECHNOLOGY
The authors declare that they have not used artificial intelligence (AI)-tools for writing and editing of the manuscript, and no images were manipulated using AI.
13. SUPPLEMENTARY MATERIAL
The supplementary material can be accessed at the journal’s website: https://jabonline.in/admin/php/uploadss/1471_pdf.pdf.
14. CONSENT TO PUBLISH
All authors have read the final manuscript and have consented to publish.
REFERENCES
1. Mustapha LS, Obayomi OV, Obayomi KS. A comprehensive review on potential heavy metals in the environment:Persistence, bioaccumulation, ecotoxicology, and agricultural impacts. Ecol Front. 2026;46:434-49. [CrossRef]
2. Kaur M, Sharma A, Aditya. A review on heavy metal accumulation and toxicity in biotic and abiotic components. IOP Conf Ser Earth Environ Sci. 2021;889:012062. [CrossRef]
3. Mir AR, Pichtel J, Hayat S. Copper:Uptake, toxicity and tolerance in plants and management of Cu-contaminated soil. Biometals. 2021;34:737-59. [CrossRef]
4. Markandeya, Mohan D, Shukla SP. Hazardous consequences of textile mill effluents on soil and their remediation approaches. Clean Eng Technol. 2022;7:100434. [CrossRef]
5. Alkhanjaf AA, Sharma S, Sharma M, Kumar R, Arora NK, Kumar B, et al. Microbial strategies for copper pollution remediation:Mechanistic insights and recent advances. Environ Pollut. 2024;346:123588. [CrossRef]
6. Orujov A, Abedi B, Wawrousek K, Aryana SA. Bio-nanocrystal stabilized biosurfactant foams for groundwater and soil remediation. InterPore J. 2025;2:260225-5. [CrossRef]
7. Araujo J, Monteiro J, Silva D, Alencar A, Silva K, Coelho L, et al. Surface-active compounds produced by microorganisms:Promising molecules for the development of antimicrobial, anti-inflammatory, and healing agents. Antibiotics (Basel). 2022;11:1106. [CrossRef]
8. Da Silva VL, Dilarri G, Mendes CR, Lovaglio RB, Gonçalves AR, Montagnolli RN, et al. Rhamnolipid from Pseudomonas aeruginosa can improve the removal of direct orange 2GL in textile dye industry effluents. J Mol Liq. 2021;321:114753. [CrossRef]
9. Hsu CY, Mahmoud ZH, Hussein UA, Abduvalieva D, Alsultany FH, Kianfar E. Biosurfactants:Properties, applications and emerging trends. S Afr J Chem Eng. 2025;53:21-39. [CrossRef]
10. Nayarisseri A, Singh P, Singh SK. Screening, isolation and characterization of biosurfactant producing Bacillus subtilis strain ANSKLAB03. Bioinformation. 2018;14:304-14. [CrossRef]
11. Biswas S, Jayaram S, Philip I, Balasubramanian B, Pappuswamy M, BarcelóD, et al. Appraisal of the potential of endophytic bacterium Bacillus amyloliquefaciens from Alternanthera philoxeroides:A triple approach to heavy metal bioremediation, diesel biodegradation, and biosurfactant production. J Environ Chem Eng. 2024;12:113454. [CrossRef]
12. Radford DS, Kihlken MA, Borrelly GP, Harwood CR, Brun NE, Cavet JS. CopZ from Bacillus subtilis interacts in vivo with a copper exporting CPx-type ATPase CopA. FEMS Microbiol Lett. 2003;220:105-12. [CrossRef]
13. Øygard JK, Måge A, Gjengedal E. Estimation of the mass-balance of selected metals in four sanitary landfills in Western Norway, with emphasis on the heavy metal content of the deposited waste and the leachate. Water Res. 2004;38:2851-8. [CrossRef]
14. Yadav AK, Kumar N, Sreekrishnan TR, Satya S, Bishnoi NR. Removal of chromium and nickel from aqueous solution in constructed wetland:Mass balance, adsorption-desorption and FTIR study. Chem Eng J. 2010;160:122-8. [CrossRef]
15. Qiao J, Borriss R, Sun K, Zhang R, Chen X, Liu Y, et al. Research advances in the identification of regulatory mechanisms of surfactin production by Bacillus:A review. Microb Cell Fact. 2024;23:100. [CrossRef]
16. Ole?ska E, Ma?ek W, Swiecicka I, Wójcik M, Thijs S, Vangronsveld J. Bacteria under metal stress-molecular mechanisms of metal tolerance. Int J Mol Sci. 2025;26:5716. [CrossRef]
17. Pratush A, Kumar A, Hu Z. Adverse effect of heavy metals (As, Pb, Hg, and Cr) on health and their bioremediation strategies:A review. Int Microbiol. 2018;21:97-106. [CrossRef]
18. Rahman FB, Sarkar B, Moni R, Rahman MS. Molecular genetics of surfactin and its effects on different sub-populations of Bacillus subtilis. Biotechnol Rep (Amst). 2021;32:e00686. [CrossRef]
19. Wróbel M, ?liwakowski W, Kowalczyk P, Kramkowski K, Dobrzy?ski J. Bioremediation of Heavy Metals by the Genus Bacillus. Int J Environ Res Public Health. 2023;20:4964. [CrossRef]
20. Tomar I, Kumari S, Joshi S, Debnath M. Harnessing the potential of bacterial consortia and biosurfactants for bioremediation of textile effluent and contaminated soil. Ind Eng Chem Res. 2024;63:14909-29. [CrossRef]
21. Sharma P, Rekhi P, Kumari S, Debnath M. Deciphering the molecular diversity of related halophilic Bacillus sp. isolated from Sambhar Lake and the functional characterizations of surfactin. J Surfactants Deterg. 2022;25(6):757–74. [CrossRef]
22. Altschul SF, Gish W, Miller W, Myers E W, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403-10. [CrossRef]
23. Fei D, Liu FF, Gang HZ, Liu JF, Yang SZ, Ye RQ, et al. A new member of the surfactin family produced by Bacillus subtilis with low toxicity on erythrocyte. Process Biochem. 2020;94:164-71. [CrossRef]
24. Lindsay WL, Norvell WA. Development of a DTPA soil test for zinc, Iron, manganese, and copper. Soil Scie Soc Am J. 1978;42:421-8. [CrossRef]
25. Rocco D, Freire BM, Oliveira TJ, Alves PL, Oliveira JM, Batista BL, et al. Bacillus subtilis as an effective tool for bioremediation of lead, copper and cadmium in water Discov Appl Sci. 2024;6:430. [CrossRef]
26. Ito K, Adachi M, Matsutani M, Kataoka R, Enomoto G, Kajikawa A, et al. Characterization of the NRPS operon homolog for surfactin A and surfactin C synthesis in Bacillus spp. Arch Microbiol. 2025;207:161. [CrossRef]
27. Selva Filho AA, Converti A, Soares Da Silva RC, Sarubbo LA. Biosurfactants as multifunctional remediation agents of environmental pollutants generated by the petroleum industry. Energies (Basel). 2023;16:1209. [CrossRef]
28. Sievers F, Wilm A, Dineen D, Gibson TJ, Karplus K, Li W, et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol Syst Biol. 2011;7:539. [CrossRef]
29. Tamura K, Stecher G, Kumar S. MEGA11:Molecular evolutionary genetics analysis version 11. Mol Biol Evol. 2021;38:3022-7. [CrossRef]
30. Jones DT, Taylor WR, Thornton JM. The rapid generation of mutation data matrices from protein sequences. Bioinformatics. 1992;8:275-82. [CrossRef]
31. Mohanty S, Jasmine J, Mukherji S. Practical considerations and challenges involved in surfactant enhanced bioremediation of oil. Biomed Res Int. 2013;2013:328608. [CrossRef]
32. Pugazhendi A, Abbad Wazin H, Qari H, Basahi JM, Godon JJ, Dhavamani J. Biodegradation of low and high molecular weight hydrocarbons in petroleum refinery wastewater by a thermophilic bacterial consortium. Environ Technol. 2017;38:2381-91. [CrossRef]
33. De Faria AF, Teodoro-Martinez DS, De Oliveira Barbosa GN, Gontijo Vaz B, Serrano Silva Í, Garcia JS, et al. Production and structural characterization of surfactin (C14/Leu7) produced by Bacillus subtilis isolate LSFM-05 grown on raw glycerol from the biodiesel industry. Process Biochem. 2011;46:1951-7. [CrossRef]
34. Rensing C, Fan B, Sharma R, Mitra B, Rosen BP. CopA:An Escherichia coli Cu(I)-translocating P-type ATPase. Proc Natl Acad Sci U S A. 2000;97:652-6. [CrossRef]
35. Ramamurthy AS, Vo D, Li XJ, Qu J. Surfactant-enhanced removal of Cu (II) and Zn (II) from a contaminated sandy soil. Water Air Soil Pollut. 2008;190:197-207. [CrossRef]
36. Dixit R, Malaviya D, Pandiyan K, Singh U, Sahu A, Shukla R, et al. Bioremediation of heavy metals from soil and aquatic environment:An overview of principles and criteria of fundamental processes. Sustainability. 2015;7:2189-212. [CrossRef]
37. Gomaa EZ, El-Meihy RM. Bacterial biosurfactant from Citrobacter freundii MG812314.1 as a bioremoval tool of heavy metals from wastewater. Bull Natl Res Cent. 2019;43:69. [CrossRef]
38. Qattan SY. Harnessing bacterial consortia for effective bioremediation:Targeted removal of heavy metals, hydrocarbons, and persistent pollutants. Environ Sci Eur. 2025;37:85. [CrossRef]
39. Olaniran AO, Balgobind A, Pillay B. Bioavailability of heavy metals in soil:Impact on microbial biodegradation of organic compounds and possible improvement strategies. Int J Mol Sci. 2013;14(5):10197-228. [CrossRef]
40. Mulligan CN, Yong RN, Gibbs BF, James S, Bennett HP. Metal removal from contaminated soil and sediments by the biosurfactant surfactin. Environ Sci Technol. 1999;33:3812-20. [CrossRef]
41. Llorca MG, Martínez-Espinosa RM. Assessment of Haloferax mediterranei genome in search of copper-molecular machinery with potential applications for bioremediation. Front Microbiol. 2022;13:895296. [CrossRef]
42. Smaldone GT, Helmann JD. CsoR regulates the copper efflux operon copZA in Bacillus subtilis. Microbiology (Reading). 2007;153:4123-8. [CrossRef]
43. Andrei A, Öztürk Y, Khalfaoui-Hassani B, Rauch J, Marckmann D, Trasnea PI, et al. Cu homeostasis in bacteria:The ins and outs. Membranes (Basel). 2020;10:242. [CrossRef]
44. Andersson M, Mattle D, Sitsel O, Klymchuk T, Nielsen AM, Møller LB, et al. Copper-transporting P-type ATPases use a unique ion-release pathway. Nat Struct Mol Biol. 2014;21:43-8. [CrossRef]
45. Kühlbrandt W. Biology, structure and mechanism of P-type ATPases. Nat Rev Mol Cell Biol. 2004;5:282-95. [CrossRef]
46. Mishra S, Lin Z, Pang S, Zhang Y, Bhatt P, Chen S. Biosurfactant is a powerful tool for the bioremediation of heavy metals from contaminated soils. J Hazard Mater. 2021;418:126253. [CrossRef]
47. Parus A, Ciesielski T, Wo?niak-Karczewska M, ?lachci?ski M, Owsianiak M, ?awniczak ?, et al. Basic principles for biosurfactant-assisted (bio)remediation of soils contaminated by heavy metals and petroleum hydrocarbons - a critical evaluation of the performance of rhamnolipids. J Hazard Mater. 2023;443:130171. CrossRef]