
INTRODUCTION
Endophytic fungi are known for their ability to stimulate plant growth by producing bioactive secondary metabolites. They are ubiquitously distributed in every plant species and significantly influence their life cycle in a symbiotic manner. Fungal endophytes also facilitate the activation of different physiological and mechanical processes of plants that help to withstand biotic and abiotic environmental stresses [1]. Triticum aestivum L. (wheat) is the second most important cereal crop for human and animal consumption. According to United Nations population predictions, India will need to produce 110–114 million tons of wheat by Mottaleb et al. [2]. In the recent era, the main concern to meet the increasing demand for staple foods is the use of chemical fertilizers on agricultural land and their long-term irreversible damage to the global ecosystem. Considering this, internal plant microorganisms with growth-promoting traits can be exclusively applied as ecofriendly inoculants for stress and disease management. Owing to its coevolutionary mode of relationship, the endophytic fungal population acts as a multilevel barrier system among various mechanisms that directly or indirectly interact to improve plant growth. Many filamentous fungal species of the genera Piriformospora [3], Aspergillus [4], Epichloe [5], Trichoderma [6], Alternaria [7], and Colletotrichum [8] have been described as plant growth promoting-endophytic fungi. Researchers have focused on studies related to the bioprospection of endophytes and understanding the potential aspects of microbial metabolites in agricultural productivity.
Indole-3-acetic acid (IAA), a crucial auxin derivative, is essential for promoting plant growth. It plays an essential role in the developmental processes of plants from embryogenesis to senescence. It also acts as a regulatory agent for cell differentiation of microbes [9]. There is evidence that IAA biosynthesis has evolved naturally in both plants and its associated micro-organisms like bacteria and fungi [10]. Several studies have demonstrated the bioactive potential of endophytic fungi in terms of environmental sustainability [11]. Furthermore, the benefits of plant growth-promoting endophytic fungi are achieved in different other ways, such as nutrient solubilization, production of volatile metabolites (e.g., HCN and ammonia), and extracellular hydrolytic enzyme production. Many studies have investigated the combined effects of exogenous auxins and endophytic metabolites on the growth characteristics of several crops, such as maize, beans, and tomatoes [12,13]. These multifaceted activities sustain plant growth and defense mechanisms [14].
Within this context, the present study was designed to isolate and identify plant growth-promoting endophytic fungi associated with an ethnomedicinal plant of Lamiaceae, Coleus forskohlii (Willd.) Briq. or “Pashanbhedi”. This is a major genus of Lamiaceae with 325 native species reported in India. It is a perennial herb with a wide range of medicinal properties, such as antidiuretic, immunosuppressant, anti-cancer, and gastro-stimulant activities. There are previous reports related to the endophytic Diaporthe, isolated from Terminalia arjuna [15], Salvia yangii [16], and Litchi chinensis [17].To the best of our knowledge, the plant growth-promoting activities of Diaporthe sp. have not been evaluated. This investigational outcome is the first report of Diaporthe osmanthi COFS1 as a potent producer of plant growth-promoting metabolites with strong in vivo utility.
2. MATERIAL AND METHODS
2.1. Collection of Plant Materials and Isolation of Endophytic Fungi
Fresh, healthy samples of C. forskohlii (Willd.) Briq. was collected from the West Midnapore district (Lat. 22.377082· N, Long. 87.040122· E), West Bengal, India, in October 2022. Labeled plants were kept in sterile ziplock bags and immediately brought to the microbiology laboratory at Vidyasagar University for endophyte isolation. Different plant parts, such as the roots, stems, and leaves, were individually surface sterilized following the procedures described by Aravind et al. [18]. A total of 75 explants were placed in water agar (2%) medium to isolate endophytes, and the plates were incubated at 30°C [19]. The fungal isolates were then transferred to PDA (Potato Dextrose Agar) containing Petri plates and kept in slants as stock cultures at 4°C.
2.2. Identification of Isolated Endophytic Fungi
Endophytic fungal isolates were identified based on macro- and micromorphological characteristics. Each isolate was observed under a Carl ZEISS Axio A1 microscope stained with lactophenol cotton blue and documented in terms of colony morphology and spore-forming structures. 5.8s rDNA-based genomic analysis was carried out for molecular identification of the fungal isolate. The aligned sequence obtained was compared with the GenBank database using BLASTn. The phylogenetic tree was constructed using MEGA 11 software following the neighbor-joining method, and sequences were submitted to the NCBI (National Center for Biotechnology Information) GenBank [20].
2.3. Screening of Fungal Endophytes for Plant Growth-Promoting (PGP) Activities
Each endophytic fungal isolate was grown in a specific medium composition following standard methods to screen for PGP attributes, such as IAA, ammonia (NH3), hydrogen cyanide (HCN), siderophore, and extracellular enzyme-producing assays. Screening for phosphate solubilization was also conducted as direct PGP activity.
Quantitative estimation of IAA production was performed using the method proposed by Patten and Glick [21]. Each fungal isolate was inoculated into CDB amended with L-tryptophan (1%) and without tryptophan, at 27°C ± 2°C for 7 days. 1 ml of both sets of supernatants was mixed with twice the volume of the Salkowaski reagent. The development of pink coloration indicated a positive response of IAA, and the concentration was estimated by measuring the absorbance at 530 nm.
The ammonia-producing ability of the endophytic isolates was estimated using the method described by Kavamura et al. [22]. Next, the HCN production was qualitatively detected using the method of Lorck [23]. Endophytic fungal isolates were transferred to PDA basal media supplemented with glycine. A Whatman, no. 1 filter paper disk saturated with 0.5% picric acid solution in 2% sodium carbonate was placed above the Petri dish. The development of a dark brown color was considered a positive result. Siderophore production was determined using Minimal Media 9 agar supplemented with Chrome Azurol S dye [24]. The change in color from blue to faint yellow was validated as siderophore-producing ability.
Fungal endophytes were screened for extracellular enzyme activity, including amylase, protease, and pectinase. Enzymatic activity was assessed by growing fungal isolates in a specific agar medium amended with 1% soluble starch [25], 1% skim milk powder [26], and 1% pectin [25]. The formation of transparent zones around fungal colonies was observed.
The qualitative phosphate solubilizing ability was determined following the method of Jasim et al. and incubated at 28°C for 4 days [27]. The solubilization index (SI) was measured using the following formula: SI (%) = (colony diameter + clear zone)/colony diameter. Quantitative estimation was performed using the National Botanical Research Institute’s Phosphate growth medium [28]. Soluble P concentrations in fungal extracts were estimated in µg ml-1.
2.4. Screening for Optimal Conditions for Auxin Production
To screen the optimal conditions for maximum IAA production, the fungal isolate was grown in different broth culture parameters, i.e., incubation time (IT) (1–10 days), pH (4.5–8.5), temperature (20°C–40°C), L-tryptophan concentration (0.1–0.7 mg ml-1), Sucrose concentration (26–34 g l-1), and NaNO3 concentration (1.0–3.0 g l-1) following the One Variable at a Time (OVAT) method. The four most significant variables, i.e., IT, medium pH (MpH), L-tryptophan concentration, and Sucrose concentration from OVAT results, were further standardized using the Box-Behnken Design (BBD) of Response Surface Methodology (RSM) with the help of Minitab software (version 21.2.0) [29]. Each independent variable was evaluated through 29 sets of experiments, with a high value denoted as “+1”, a low value denoted as “-1”, and an optimum value marked as “0”. The predicted responses were calculated using a regression equation, and the interactions of the effective variables were represented as surface plots.
2.5. Characterization of Fungal Metabolites
Endophytic fungal isolate was cultured in large-scale fermentation, and the cell-free culture filtrate was adjusted to pH 2.8 with 1N HCL and extracted with twice the volume of ethyl acetate (1:2). Solvent extraction was used to extract crude metabolites from D. osmanthi COFS1. The ethyl acetate extract was examined under 200–400 nm of wavelength in Labman LMSPUV1200 spectrophotometer. Standard IAA and crude extract were then eluted using isopropanol:n-butanol:ammonia:water (50:30:15:5) solvent system [30]. Silica-gel-coated alumina plates were visualized under 254 nm UV light. The presence of different functional groups within the crude extract was analyzed using an FT-IR (PerkinElmer FT-IR C98747) spectrophotometer in the 400–4,000 cm-1 wave range.
Visual bands were scraped from silica-coated plates, re-dissolved in methanol, and analyzed. Metabolites produced by COFS1 in culture broth were determined using Gas Chromatography (TRACE 1300) Mass Spectrometry (ISQ QD Single Quadrupole) system (GC-MS) from Thermo Scientific in ESI mode [31]. According to Bhanja and Rout [32], the methanolic extract was run in an Agilent 1100 liquid chromatography-quantitative-time of flight (LC-Q-TOF) system using an isocratic mixture of 0.1% formic acid and acetonitrile. Nuclear magnetic resonance (NMR) analysis of purified COFS1 extract was performed at Vidyasagar University Science Instrumentation Center using a Brucker-Avance DPX FT-NMR 400 MHz spectrometer (Brucker Corporations, USA). Chemical shifts of the metabolite are documented in parts per million using the DMSO-d6 solvent system.
2.6. Plant Growth-Promoting Effects of Cell-Free Culture Extracts
To assess growth-promoting activity, wheat (Triticum aestivum L. “PBW343”) was used as a model plant. The potent endophytic isolate D. osmanthi COFS1 was cultured in CDB for 10 days at 27°C with 150 rpm. The culture filtrate was then centrifuged at 10,000 rpm for 15 minutes at 4°C, followed by filtration using a Whatman filter paper (Grade 1:11 µm). The supernatant was used as an elicitor and diluted with sterile distilled water to 20%, 50%, and 80%. Seeds were sterilized with 1% sodium hypochlorite solution for 10 minutes and 75% ethanol for 1 minute and rinsed twice with ddH2O. After disinfection, seeds were germinated in three different concentrations of culture filter within 9-cm Petri dishes containing filter paper. Petri plates containing sterile distilled water and CDB were used as control plates. After the emergence of radicals and plumules, three seedlings from each treatment were transferred to sterilized plastic bags containing autoclaved soil and coco-peat mixture (1:1). At the two-leaf stage, 20 ml of each extract concentration was applied to the apex at an interval of 2 days. Various growth attributes, such as fresh and dry weight (in gm), shoot and root length (in cm), chlorophyll content, and carotenoid content were observed after 60 days of treatment.
2 .7. Statistical Analysis
Data obtained from several experiments were performed in triplicate and analyzed using one-way analysis of variance with GraphPad Prism 8.4.2 software (La Jolla, CA, USA). Differences in experimental outcomes were compared at p < 0.05 probability values using Tukey’s multiple comparison test.
3. RESULTS
3.1. Isolation and Identification of Fungal Endophytes
A total of 13 endophytic fungal genera were isolated from different plant parts of C. forskohlii; including three isolates from roots, five from stems, and five from leaves. These isolates were initially differentiated on the basis of macro- and micromorphological characteristics and identified as several fungal genera, of which Fusarium was the only genus associated with each plant tissue. Stem tissues were harbored by the maximum number of fungal isolates, with a colonization rate of 42% (Supplementary Table 1). The potent isolate COFS1 was subsequently subjected to 5.8s rDNA sequencing, which revealed it to be more than 99.60% identical to Diaporthe osmanthi (GenBank accession: ON927178) (Fig. 1).
3.2. Screening of Fungal Endophytes for PGP-Attributes
Out of the 13 endophytes, 6 fungal isolates exhibited IAA production activity (0.21–13.12 µg ml-1) without tryptophan, while 10 fungal isolates were able to synthesize auxin at various concentrations (0.18–31.72 µg ml-1) in broth amended with 0.1% L-tryptophan. The isolate D. osmanthi COFS1 produced the highest concentration of IAA regardless of the presence of a precursor (Fig. 2 and Supplementary Table 2). The production of ammonia, HCN, and siderophores was positive for three, one, and two fungal strains, respectively. Three fungal isolates (i.e., 23%) tested positive for amylase activity, one isolate (i.e., 7%) for protease activity, and two isolate (i.e., 15%) for pectinase activity. The endophytic isolates COFR2 (SI:1.14 ± 0.01) and COFS1 (SI:1.70 ± 0.01) showed PGP attributes of phosphate solubilization (Table 1). On the fifth day of incubation, with a pH of 5.2, the maximum solubilization (31.85 µg ml-1) of phosphate by the COFS1 isolate was observed (Fig. 3). Amylase enzyme solubilization index of this endophytic isolate was 0.34, whereas 1.62 was for the pectinase enzyme.
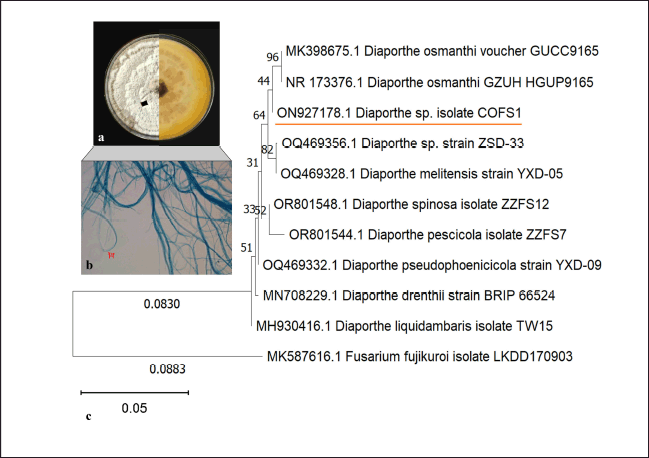 | Figure 1. Plate morphology and microscopic characteristics of isolate COFS1 (a,b); Phylogenetic tree of the isolate constructed using the neighbor-joining method with bootstrap values (c). [Click here to view] |
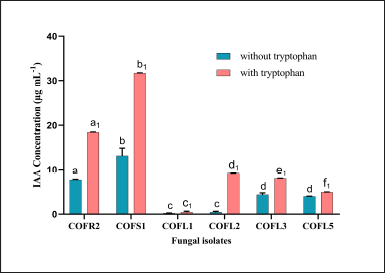 | Figure 2. IAA production by endophytic isolates with and without L-tryptophan. Different letters (i.e., a1-f1 for IAA production with tryptophan and a-d for IAA production without tryptophan) indicate valid statistical differences, whereas a similar letter indicates no difference (p < 0.05). [Click here to view] |
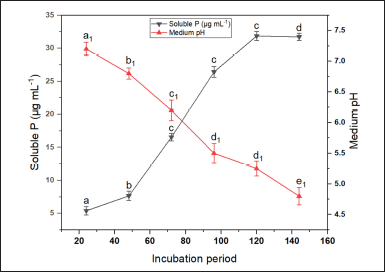 | Figure 3. Phosphate solubilizing profile of COFS1 in response to medium pH and IT. The values in the graphs are mean ± standard error of three replicates. A different letter (a–d for soluble phosphate and a1–e1 for medium pH) indicates a valid statistical difference, whereas a similar letter indicates no such difference. (p < 0.05). [Click here to view] |
3.3. Optimization of the Growth Parameters
Auxin production started to increase after 24 hours of incubation, reaching its maximum (31.72 µg ml-1) on the seventh day of the fermentation period (Fig. 4a). Medium pH 6.5 significantly influenced IAA production, reaching up to 32.02 µg ml-1, with a sharp decline observed at alkaline pH values (Fig. 4b). However, the temperature range between 25°C and 35°C was positively influenced metabolite production in broth (Fig. 4c). On the other hand, a steep increase was noticed between 0.1 and 0.5-mg ml-1 of L-tryptophan supplemented conditions (Fig. 4d). The least amounts of fungal growth and IAA were obtained when the fermentation broth was supplemented with maltose. Furthermore, NaNO3 (19.22 µg ml-1) followed by KNO3 (7.77 µg ml-1) was effective in comparison with other nitrogen sources. Similarly, while optimizing different concentrations of both carbon and nitrogen sources, 30 g l-1 sucrose (35.63 µg ml-1) and 2.0 g l-1 NaNO3 (28.63 µg ml-1) showed maximum influence on auxin production (Fig. 4e and f). The OVAT results revealed that fermentation conditions at MpH 6.5, growth temperature 30°C, amended with 0.5 mg ml-1 of L-tryptophan, 30 g l-1 of sucrose, and, 2 g l-1 of NaNO3 on the seventh day of incubation were optimum for the highest IAA production (Supplementary table 3).
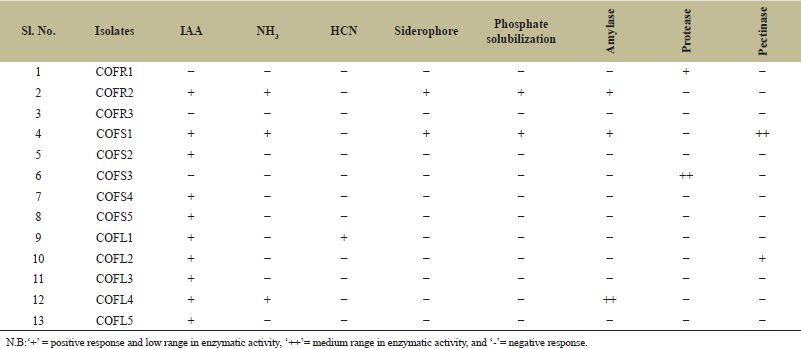 | Table 1. PGP activities of the endophytic fungal isolates of C. forskohlii. [Click here to view] |
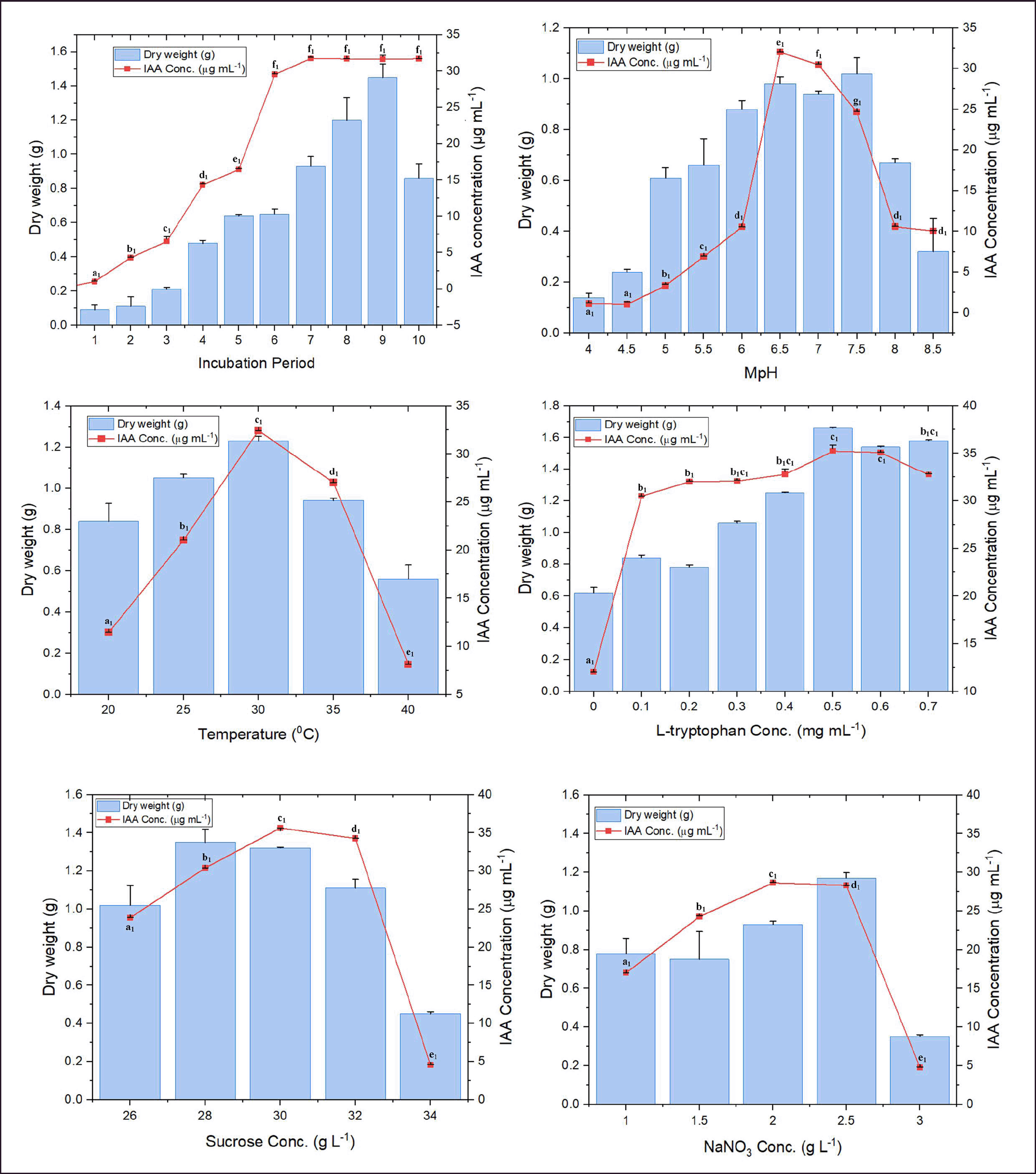 | Figure 4. Influence of different culture parameters on the IAA production and growth (dry weight of fungal biomass) of D. osmanthi COFS1. Values represent the mean ± standard error of three replicates. A different letter (a1–e1) indicates a valid statistical difference, whereas a similar letter indicates no such difference. (p < 0.05). [Click here to view] |
The four most effective fermentation parameters, namely, IT, medium pH, L-tryptophan concentration, and sucrose concentration, were further analyzed using RSM. According to BBD, the fungal isolate COFS1 showed the highest IAA production of 37.30 µg ml-1 at pH 6.5, 30 g l-1 of sucrose, and media amended with 0.5 mg ml-1 of L-tryptophan at seventh days of incubation. Whereas, the least amount of IAA produced, i.e., 16.73 µg ml-1, when media supplemented with 0.4 mg ml-1 of L-tryptophan, 30 g l-1 of sucrose, 6.5 pH on the sixth day of IT (Table 2). The model F-value (8756.37) was significant, whereas model p-values (p < 0.001) were highly significant, with a 0.00% chance of error. A predicted R2 of 99.95% agreed with the adjusted R2 of 99.98%, and optimum values of independent variables that affected the response were evaluated using 3D surface plots (Fig. 5a1-f1 and Supplementary Table 4). There is no significant difference between the actual response (37.44 µg ml-1) of the laboratory experiment and the predicted response (37.37 µg ml-1) of RSM adopting a fermentation condition of 7 incubation days, 6.5 pH, supplemented with 0.51 mg ml-1 L-tryptophan and 30.15 g l-1of sucrose.
3.4. Characterization of PGP Metabolites
UV-VIS spectral peak was obtained at λmax275.51 nm. The crude extract of the fungal isolate exhibited three bands with Rf values of 0.9, 0.89, and 0.84. The Rf value of band 2 was similar to that of standard IAA (Supplementary Figure 1a and b). The IR spectrum of COFS1 extracts at 2,990 cm-1 is attributed to the presence of alkyl (-CH2) stretching. The peak at 1,740 cm-1 is ascribed to the C=O bond of the indole. The absorption peaks at 1,240 and 1,092 cm-1 are attributed to C-N stretching. Furthermore, wagging at 930, 844, and 610 cm-1 indicated C-H vibration associated with the indole ring structure (Supplementary Figure 2).
To identify the indole compounds produced by the COFS1 isolate, the culture filtrate was analyzed by GC-MS. In total, nine major compounds were identified at different retention times (Supplementary Table 5). The majority of these compounds belong to indole groups, such as 1-methylene-1H-indene, 1H-Indole, Indole-3-acetic acid, Tryptophol, Indole-6-carboxaldehyde, and 2H-Benzopyran-2-one. IAA in the culture filtrate was detected at a retention time of 14.50 minutes with an area percentage of 40.99 (Fig. 6). The methanolic fraction of band 2 from the TLC plate was then analyzed using HRMS, which showed different absorption peaks in the m/z range of 173.01-212.06 (Supplementary Figure 3). The H1-NMR spectrum showed peaks at δ ppm, which were 7.521–7.559 (m, 2H), 7.312–7.368 (m, 1H), 7.114–7.117 (d, 1H), 7.001–7.097 (m, 2H), 6.923–6.985 (m, 1H), and 3.502 (s, 1H) (Fig. 7).
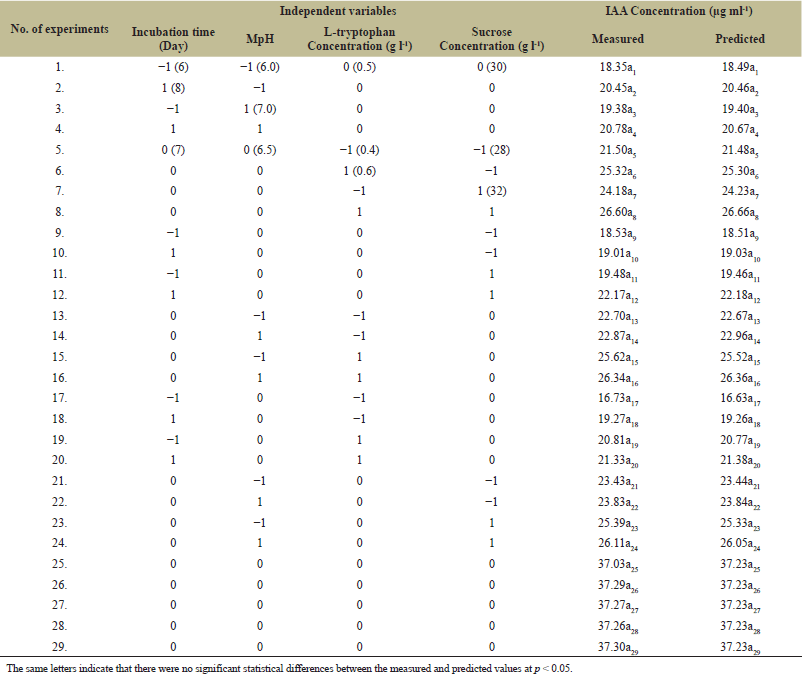 | Table 2. Experimental design and BBD results for optimizing IAA production from COFS1. [Click here to view] |
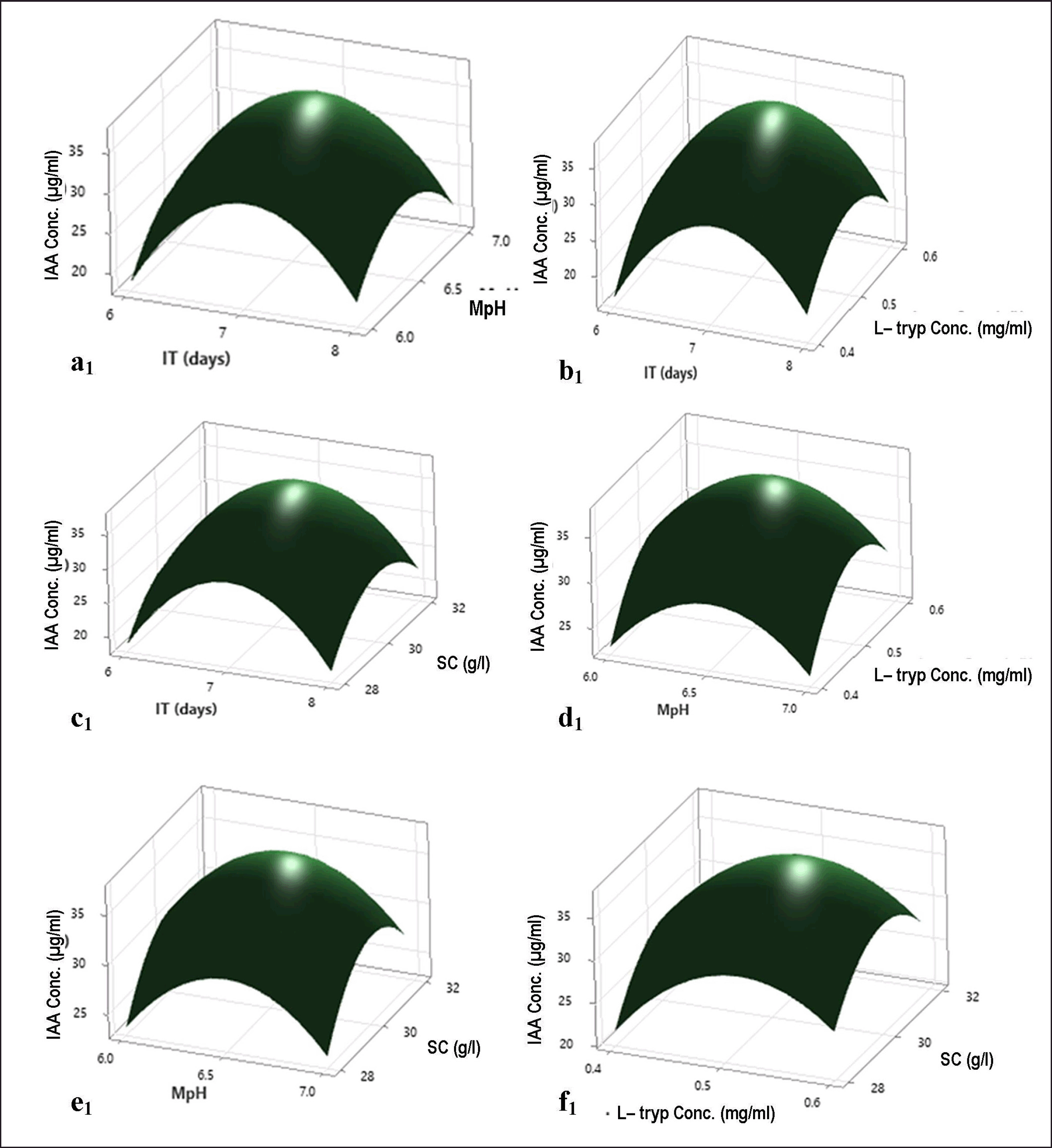 | Figure 5. Response surface plots demonstrating the effects of different media compositions on IAA production by COFS1; (a1) interaction between IT (days) versus MpH, (b1) interaction between L-tryp Conc. (mg/ml) versus IT (days), (c1) interaction between SC (g/l) versus IT (days), (d1) interaction between L-tryp Conc. (mg/ml) versus MpH, (e1) interaction between MpH vs. SC (g/l),and (f1) interaction between SC (g/l) vs. L-tryp Conc. (mg/ml). [Click here to view] |
3.5. Evaluation of Growth-Promoting Attributes
Wheat (Triticum aestivum L.) seedlings treated with different concentrations of fungal extract enhanced growth attributes significantly. Each experiment was conducted in triplicate. Each plant was harvested after 60 days of treatment, and the growth parameters were measured (Supplementary Figure 4). Under the experimental conditions, we observed a statistically significant difference between the seedlings treated with 20%, 50%, and 80% fungal extracts and the control group. Maximum growth attributes were recorded at 50% diluted fermentation broth, like fresh weight (4.31 ± 0.20b), dry weight (1.12 ± 0.24b), shoot length (36.20 ± 0.55b), root length (28.30 ± 0.11c), and chlorophyll content (3.61 ± 0.12b). Plants treated with 50% fungal extract showed increased fresh weight (1.35-fold), dry weight (2.48-fold), shoot length (1.02-fold), root length (1.40-fold), chlorophyll content (1.55-fold), and carotenoid content (1.82-fold), respectively, compared with the sterile water- and CDB-treated plants (Fig. 8). These results indicate that the culture filtrate of the endophytic COFS1 isolate promotes wheat growth under in vivo conditions.
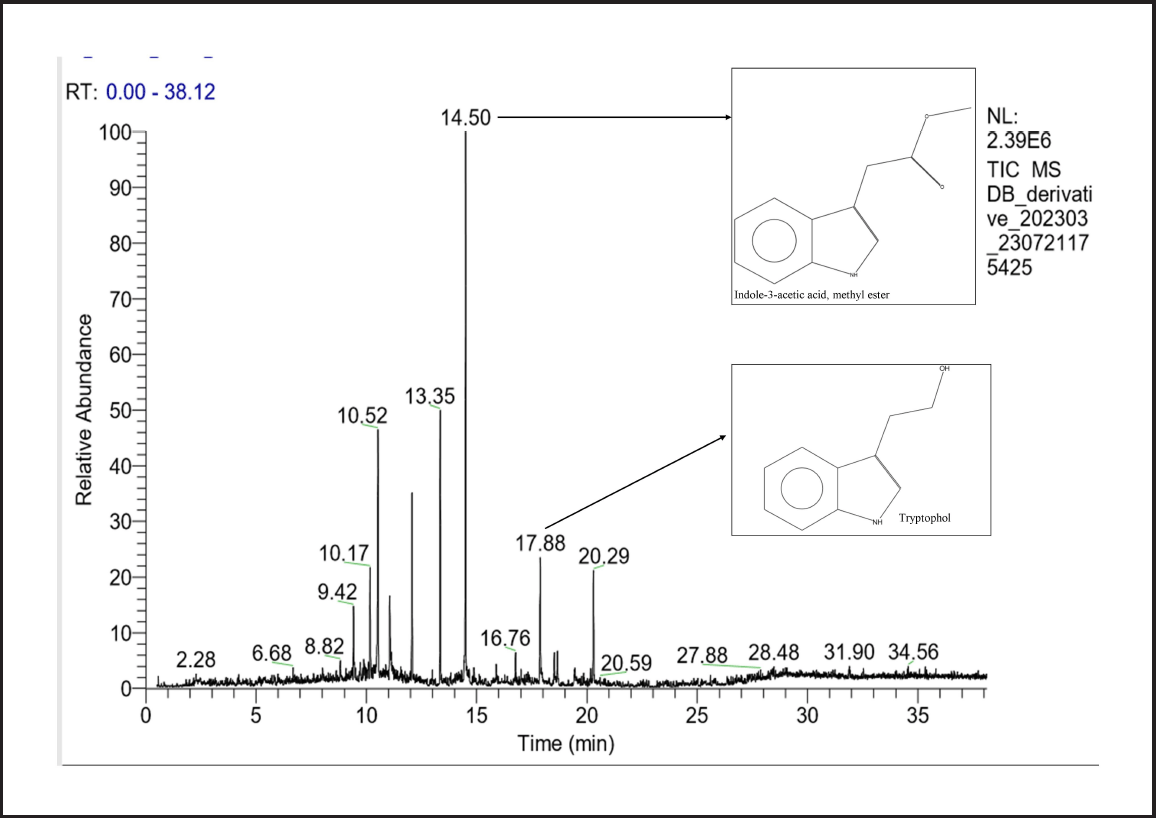 | Figure 6. Growth-promoting metabolites produced by D. osmanthi COFS1 identified through GC-MS (NIST Library). [Click here to view] |
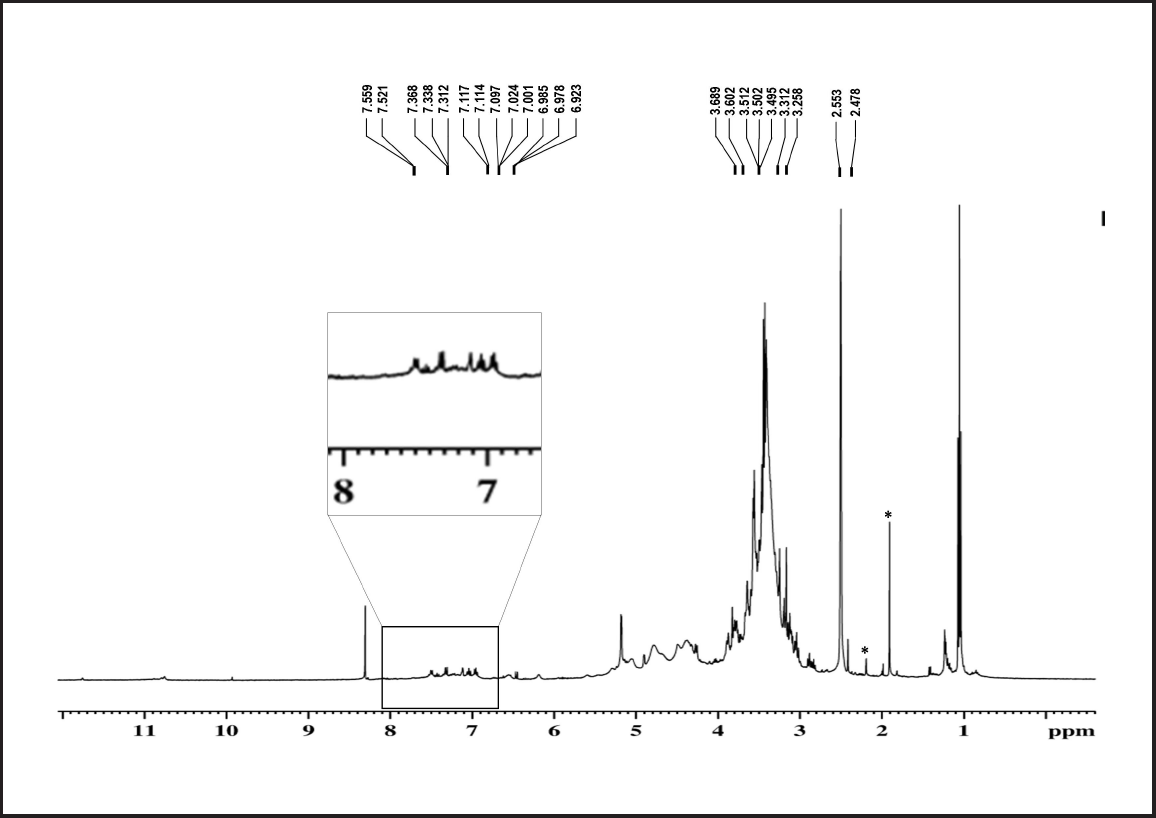 | Figure 7. H1-NMR spectrum of band 2 of the COFS1 isolate (‘*’ is representing as solvent peaks). [Click here to view] |
4. DISCUSSION
The ubiquitous distribution of endophytic fungi, irrespective of the host tissue, represents them as beneficial sources of bioactive metabolites. It has been estimated that over a million endophytic fungal species occupy every ecosystem and defend their host plants [33]. With the recent advancement in plant-endophyte interaction, one of the interesting benefits is the promotion of plant growth. Endophytic fungi trigger plant growth-promoting mechanisms in response to environmental stimuli. Numerous studies have reported on the growth-promoting endophytic-fungal community associated with different medicinal plants worldwide [34]. Earlier studies reported that endophytic fungi, such as Alternaria alternata, Aspergillus niger, and Trichoderma harzianum, enhance wheat production through phosphate solubilization, nitrogen transfer, IAA, and siderophore production [6,35]. The endophytic fungal diversity associated with C. forskohlii has been reported by Crasta and Raveesha [36], which signifies the presence of 34 fungal genera, of which the majority belongs to Ascomycota. This experimental study highlighted the PGP activity of the endophytic strain D. osmanthi, which has been identified as a potent plant growth inducer.
 | Figure 8. Effects of different dilutions of the culture extract of the fungal endophyte D. osmanthi COFS1 on the growth attributes of wheat plants. [Click here to view] |
In this study, a total of 13 endophytic fungal isolates from C. forkohlii were screened for PGP activity. Various fungal genera, including Fusarium, Cochliobolus, Nigrospora, Cladosporium, Helminthosporium, and Chrysosporium, were found in specific plant parts, confirming the tissue-specific occurrence of endophytes. The most potent isolate, D. osmanthi COFS1, showed promising auxin-producing ability (31.72 µg ml-1) in the presence of L-tryptophan. Fungal IAA along with host endogenous auxin affects plant cell division and differentiation and stimulates vascular tissue and root development [37]. On the other hand, solubilization of limiting nutrients like phosphorus directly enhances the mineral mobility toward root tissues. In our study, the highest P-solubilization activity was also exhibited by the fungal isolate D. osmanthi COFS1 (31.85 µg ml-1) related to a gradual decrease in pH due to organic acid production. This isolate could produce NH3 and siderophores under the experimental conditions. Phosphate availability helps increase the photosynthetic rate and lateral root formation, and siderophores accelerate plant growth under iron-limiting conditions [38]. A recent study by Toghueo et al. [39] also reported the IAA and siderophore production in the endophytic Diaporthe strain EB4. Thus, the multidimensional PGP activities of endophytic fungi play an important role in achieving overall crop yields and developmental processes.
To maximize the fungal metabolite production during large-scale processes, it is necessary to optimize fermentation parameters. In this study, the OVAT analysis, coupled with a response surface model, provided a clear understanding of the interactions among different media. Previous studies have discussed the effects of fermentation conditions on higher IAA yields [40]. The optimization result of this study revealed that maximum IAA production was recorded on the seventh day of the incubation period at a media temperature of 30°C, pH 6.5, L-tryptophan concentration of 0.5 mg ml-1, sucrose concentration of 30 g l-1, and NaNO3 concentration 2 g l-1. This is in accordance with the reports of Numponsak et al. [8] and Jagannath et al. [41]. There was a sharp decrease in IAA production after a certain growth parameter range. These findings agree with those of Bar and Okon [42], who reported that a higher concentration of L-tryptophan has a negative effect on microbial growth and actively affects IAA breakdown. Auxin production increased approximately 1.30-fold by adopting response surface model values.
The ethyl acetate extracts from the fungal isolate showed a peak at λmax275.51 nm, which is consistent with the results of Kaneshiro et al. [43]. The Rf value for the COFS1 extract enriched with IAA was 0.89, which was consistent with the standard IAA. These outcomes are similar to those of Harikrishnan et al. [44]. The FT-IR spectrum confirmed the presence of functional groups associated with the indole structure, which is in accordance with the findings of Jaganath et al. [41]. The GC-MS chromatogram indicated several indole compounds, of which the methyl ester of indole-3-acetic acid was the dominant compound with a greater area percentage. This result is consistent with previous studies that explained the protective role of IAA conjugates in auxin degradation in plants [45]. Along with this, tryptophol obtained from the extract suggested an IAA biosynthesis pathway through IPA (Indole-3-pyruvic acid) [46]. Data obtained from the HRMS analysis bear resemblance with the fragmentation patterns of the indole compounds at 174.98 and 212.06 m/z ratio [47]. Furthermore, 1H NMR showed peaks at 2.478–2.553 and 6.923–7.559 ppm characterized by the side chain of –CH2 and indole ring protons [41,48]. Our findings indicate that the endophytic fungal isolate D. osmanthi COFS1 can synthesize IAA and other growth-promoting metabolites to accelerate overall plant development.
Previous studies have shown that an increase in the length of plant roots and shoots was a positive indicator of adequate growth [49]. The positive effects of fungal culture extract on wheat growth attributes were demonstrated under in vivo experimental conditions. An increase in the fresh and dry weights of plants was observed when 50% of the fungal extract was applied compared with the uninoculated controls. Among different concentrations of fungal extract, a 50% dilution of culture extract displayed a considerable increase in terms of shoot-root length, leading to a 1.02-fold increase in shoot length, and a 1.40-fold increase was observed in root lengths. The significant increase in pigment content, such as chlorophyll and carotenoids, further demonstrated the growth-inducing ability of the isolates. In a similar study, Khan et al. [50] demonstrated the PGP abilities of the endophytic fungus Acremonium sp. and the effect of fermentation broth on the growth of Allium tuberosum. Mohamed et al. [51] also highlighted the plant growth-promoting traits of the endophytic fungus Fusarium petersiae on the overall plant growth of wheat seedlings under low nitrogen availability. Higher plant biomass production is directly related to the up-regulation of several structural genes, which facilitate proper penetration of endophytic fungi and enhance root length, along with an increase in photosynthetic rate [52,53]. D. masirevici and D. terebinthifolii have been studied for IAA production [54,55]. In addition, the endophytic fungus Colletotrichum fructicola CMU-A109 and an actinobacterial species produce potent IAA production [8,56]. This study reports the ability of the fungal isolate D. osmanthi COFS1 to produce IAA, ammonia, siderophore, and growth-inducing secondary metabolites, thereby recognizing this endophyte as a potential plant-growth stimulator.
5. CONCLUSION
This study represents the first report on the growth-promoting effects of endophytic fungi isolated from C. forskohlii. Among these isolates, D. osmanthi COFS1 had the best potential for nutrient solubilization, IAA, and siderophore production as well as the synthesis of hydrolytic enzymes; hence, it was selected for bioassays on wheat seedlings. The enhanced growth parameters along with the presence of several bioactive metabolites in the crude ethyl acetate extract of this fungal isolate strongly indicate that this endophytic strain possesses considerable plant growth-promoting activities. These findings suggest that D. osmanthi COFS1 should be further explored as an effective bioinoculant for the sustainable cultivation of other cash crops.
6. ACKNOWLEDGMENTS
DB and RD would like to thank the University Sophisticated Instrument Center (USIC) of Vidyasagar University. The authors are grateful to the Microbiology and Microbial Biotechnology Laboratory of the Department of Botany and Forestry, Vidyasagar University, for their support and access to the required facilities.
7. CONFLICTS OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
8. FUNDING SOURCES
There is no funding to report.
9. AUTHOR CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work. All the authors are eligible to be an author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
10. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
11. DATA AVAILABILITY
All datasets interpreted and analyzed in this study are included in the manuscript and/or supplementary files.
12. PUBLISHER’S NOTE
All claims expressed in this article are solely those of the authors and do not necessarily represent those of the publisher, the editors and the reviewers. This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
13. USE OF ARTIFICIAL INTELLIGENCE (AI)-ASSISTED TECHNOLOGY
The authors declares that they have not used artificial intelligence (AI)-tools for writing and editing of the manuscript, and no images were manipulated using AI.
REFERENCES
1. Kumar A, Verma JP. Does plant—microbe interaction confer stress tolerance in plants: a review?.Microbiol Res 2018; 207:41–52; CrossRef
2. Mottaleb KA, Kruseman G, Frija A, Sonder K, Lopez-Ridaura S. Projecting wheat demand in China and India for 2030 and 2050: implications for food security. Front Nutr 2023;26(9):1077443; CrossRef
3. Sabra M, Aboulnasr A, Franken P, Perreca E, Wright LP, Camehl I. Beneficial root endophytic fungi increase the growth and quality parameters of sweet basil in heavy metal contaminated soil. Front Plant Sci 2018;9:1726; CrossRef
4. Mehmood A, Hussain A, Irshad M, Hamayun M, Iqbal A, Khan N. In vitro production of IAA by endophytic fungus Aspergillus awamori and its growth promoting activities in Zea mays. Symbiosis 2019;77:225–35; CrossRef
5. Vignale MV, Iannone LJ, Scervino JM, Novas MV. Epichloë exudates promote in vitro and in vivo arbuscular mycorrhizal fungi development and plant growth. Plant Soil 2018;422:267–81; CrossRef
6. Ripa FA, Cao WD, Tong S, Sun JG. Assessment of plant growth promoting and abiotic stress tolerance properties of wheat endophytic fungi. Biomed Res Int 2019;2019(1):6105865; CrossRef
7. Khan AR, Ullah I, Waqas M, Shahzad R, Hong SJ, Park GS, et al. Plant growth-promoting potential of endophytic fungi isolated from Solanum nigrum leaves. World J Microbiol Biotechnol 2015;31:1461–6; CrossRef
8. Numponsak T, Kumla J, Suwannarach N, Matsui K, Lumyong S. Biosynthetic pathway and optimal conditions for the production of indole-3-acetic acid by an endophytic fungus, Colletotrichum fructicola CMU-A109. PLOS One 2018;13(10):e0205070; CrossRef
9. Yashavantha Rao HC, Santosh P, Rakshith D, Satish S. Molecular characterization of an endophytic Phomopsis liquidambaris CBR-15 from Cryptolepisbuchanani Roem. and impact of culture media on biosynthesis of antimicrobial metabolites. 3 Biotech 2015;5:165–73; CrossRef
10. Fu SF, Wei JY, Chen HW, Liu YY, Lu HY, Chou JY. Indole-3-acetic acid: a widespread physiological code in interactions of fungi with other organisms. Plant Signal Behav 2015;10(8):e1048052; CrossRef
11. Anand U, Pal T, Yadav N, Singh VK, Tripathi V, Choudhary KK, et al. Current scenario and future prospects of endophytic microbes: promising candidates for abiotic and biotic stress management for agricultural and environmental sustainability. Microbial ecol 2023;86(3):1455–86; CrossRef
12. Baron NC, de Souza Pollo A, Rigobelo EC. Purpureocilliumlilacinum and Metarhizium marquandii as plant growth-promoting fungi. PeerJ. 2020;8:e9005; CrossRef
13. Ismail MA, Amin MA, Eid AM, Hassan SE, Mahgoub HA, Lashin I, et al. Comparative study between exogenously applied plant growth hormones versus metabolites of microbial endophytes as plant growth-promoting for Phaseolus vulgaris L. Cells. 2021;10(5):1059; CrossRef
14. Vinayarani G, Madhusudhan KN, Prakash HS. Induction of systemic resistance in turmeric by rhizospheric isolate Trichoderma asperellum against rhizome rot disease. J Plant Pathol 2019;101:965–80; CrossRef
15. Patil M, Patil R, Mohammad S, Maheshwari V. Bioactivities of phenolics-rich fraction from Diaporthearengae TATW2, an endophytic fungus from Terminalia arjuna (Roxb.). Biocatal Agric Biotechnol 2017;10:396–402; CrossRef
16. Zimowska B, Bielecka M, Abramczyk B, Nicoletti R. Bioactive products from endophytic fungi of sages (Salvia spp.). Agriculture 2020;10(11):543; CrossRef
17. Huang S, Xia J, Zhang X, Sun W. Morphological and phylogenetic analyses reveal three new species of Diaporthe from Yunnan, China. MycoKeys 2021;78:49; CrossRef
18. Aravind R, Kumar A, Eapen SJ, Ramana KV. Endophytic bacterial flora in root and stem tissues of black pepper (Piper nigrum L.) genotype: isolation, identification and evaluation against Phytophthora capsici. Lett applied microbiol 2009;48(1):58–64; CrossRef
19. Schulz B, Boyle C, Draeger S, Römmert AK, Krohn K. Endophytic fungi: a source of novel biologically active secondary metabolites. Mycological Res 2002;106(9):996–1004; CrossRef
20. Tamura K, Dudley J, Nei M, Kumar S. MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol Biol Evol 2007;24(8):1596–9; CrossRef
21. Patten CL, Glick BR. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl Environ Microbiol 2002;68(8):3795–801; CrossRef
22. Kavamura VN, Santos SN, da Silva JL, Parma MM, Ávila LA, Visconti A, et al. Screening of Brazilian cacti rhizobacteria for plant growth promotion under drought. Microbiol Res 2013;168(4):183–91; CrossRef
23. Lorck H. Production of hydrocyanic acid by bacteria. Physiol Plant 1948;1(2):142–6.
24. Pérez-Miranda S, Cabirol N, George-Téllez R, Zamudio-Rivera LS, Fernández FJ. O-CAS, a fast and universal method for siderophore detection J Microbiol Methods 2007;70(1):127–31; CrossRef
25. Sunitha VH, Devi DN, Srinivas C. Extracellular enzymatic activity of endophytic fungal strains isolated from medicinal plants. World J Agric Sci 2013;9(1):01–9; CrossRef
26. Amirita A, Sindhu P, Swetha J, Vasanthi NS, Kannan KP. Enumeration of endophytic fungi from medicinal plants and screening of extracellular enzymes. World J Sci Technol 2012;2(2):13–9.
27. Jasim B, Joseph AA, John CJ, Mathew J, Radhakrishnan EK. Isolation and characterization of plant growth promoting endophytic bacteria from the rhizome of Zingiber officinale. 3 Biotech 2014;4:197–204; CrossRef
28. Nautiyal CS. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol Lett 1999;170(1):265–70; CrossRef
29. Box GE, Behnken DW. Some new three level designs for the study of quantitative variables. Technometrics 1960;2(4):455–75.
30. Goswami D, Pithwa S, Dhandhukia P, Thakker JN. Delineating Kocuriaturfanensis 2M4 as a credible PGPR: a novel IAA-producing bacteria isolated from saline desert. J Plant Interact 2014;9(1):566–76; CrossRef
31. Shahzad R, Waqas M, Khan AL, Al-Hosni K, Kang SM, Seo CW, et al. Indoleacetic acid production and plant growth promoting potential of bacterial endophytes isolated from rice (Oryza sativa L.) seeds. Acta Biol Hung 2017;68(2):175–86; CrossRef
32. Bhanja SK, Rout D. Structural analysis of two bioactive components of an Edible Mushroom, Termitomyces microcarpus. Nat Prod Commun 2017;12(12):1934578X1701201226.
33. Lugtenberg BJ, Caradus JR, Johnson LJ. Fungal endophytes for sustainable crop production. FEMS Microbiol Ecol 2016;92(12):fiw194; CrossRef
34. Rana KL, Kour D, Kaur T, Devi R, Yadav AN, Yadav N, et al. Endophytic microbes: biodiversity, plant growth-promoting mechanisms and potential applications for agricultural sustainability. Antonie Van Leeuwenhoek 2020;113:1075–107; CrossRef
35. Qiang X, Ding J, Lin W, Li Q, Xu C, Zheng Q, et al. Alleviation of the detrimental effect of water deficit on wheat (Triticum aestivum L.) growth by an indole acetic acid-producing endophytic fungus. Plant Soil 2019;439:373–91; CrossRef
36. Crasta GL, Raveesha KA. Molecular identification of endophytic fungi associated with Coleus forskohlii (Willd.) Briq. J Appl Biol Biotech 2021;9(06):162–72.
37. Kumar VV. Plant growth-promoting microorganisms: interaction with plants and soil. In: Hakeem K, Akhtar M, Abdullah S (eds.). Plant, soil and microbes. Springer, Cham, Switzerland, vol. 1, pp 1–6, 2016; CrossRef
38. Pérez-Torres CA, Lopez-Bucio J, Cruz-Ramírez A, Ibarra-Laclette E, Dharmasiri S, Estelle M, et al. Phosphate availability alters lateral root development in Arabidopsis by modulating auxin sensitivity via a mechanism involving the TIR1 auxin receptor. Plant Cell 2008;20(12):3258–72; CrossRef
39. Toghueo RM, Vázquez de Aldana BR, Zabalgogeazcoa I. Diaporthe species associated with the maritime grass Festuca rubra subsp. pruinosa. Front Microbiol 2023;14:1105299; CrossRef
40. Galeano RM, Franco DG, Chaves PO, Giannesi GC, Masui DC, Ruller R, et al. Plant growth promoting potential of endophytic Aspergillus niger 9-p isolated from native forage grass in Pantanal of Nhecolândia region, Brazil. Rhizosphere 2021;18:100332; CrossRef
41. Jagannath S, Konappa NM, Alurappa R, Chowdappa S. Production, characterization of indole acetic acid and its bioactive potential from endophytic fungi of Cymbidium aloifolium L. J Biol Active Prod Nat 2019;9(5):387–409; CrossRef
42. Bar T, Okon Y. Tryptophan conversion to indole-3-acetic acid via indole-3-acetamide in Azospirillumbrasilense Sp7. Canadian J Microbiol 1993;39(1):81–6; CrossRef
43. Kaneshiro T, Slodki ME, Plattner RD. Tryptophan catabolism to indole pyruvic and indole acetic acids by Rhizobium japonicum L-259 mutants. Curr Microbiol 1983;8:301–6; CrossRef
44. Harikrishnan H, Shanmugaiah V, Balasubramanian N, Sharma MP, Kotchoni SO. Antagonistic potential of native strain Streptomyces aurantiogriseus VSMGT1014 against sheath blight of rice disease. World J Microbiol Technol 2014;30:3149–61; CrossRef
45. Nutaratat P, Srisuk N, Arunrattiyakorn P, Limtong S. Indole-3-acetic acid biosynthetic pathways in the basidiomycetous yeast Rhodosporidiumpaludigenum. Arch Microbiol 2016;198:429–37; CrossRef
46. Bunsangiam S, Sakpuntoon V, Srisuk N, Ohashi T, Fujiyama K, Limtong S. Biosynthetic pathway of indole-3-acetic acid in basidiomycetous yeast Rhodosporidiobolusfluvialis. Mycobiol 2019;47(3):292–300; CrossRef
47. Revelou PK, Kokotou MG, Constantinou-Kokotou V. Identification of auxin metabolites in Brassicaceae by ultra-performance liquid chromatography coupled with high-resolution mass spectrometry. Molecules 2019;24(14):2615; CrossRef
48. Zhang BX, Li PS, Wang YY, Wang JJ, Liu XL, Wang XY, et al. Characterization and synthesis of indole-3-acetic acid in plant growth promoting Enterobacter sp. RSC Adv 2021;11(50):31601–7; CrossRef
49. Wang Y, Thorup-Kristensen K, Jensen LS, Magid J. Vigorous root growth is a better indicator of early nutrient uptake than root hair traits in spring wheat grown under low fertility. Front Plant Sci 2016;7:865; CrossRef
50. Khan MS, Gao J, Munir I, Zhang M, Liu Y, Moe TS, et al. Characterization of endophytic fungi, Acremonium sp., from Lilium davidii and analysis of its antifungal and plant growth-promoting effects. BioMed Res Int 2021;2021(1):9930210; CrossRef
51. Mohamed AH, Abd El-Megeed FH, Hassanein NM, Youseif SH, Farag PF, Saleh SA, et al. Native rhizospheric and endophytic fungi as sustainable sources of plant growth promoting traits to improve wheat growth under low nitrogen input. J Fungi 2022;8(2):94; CrossRef
52. Harman GE, Uphoff N. Symbiotic root- endophytic soil microbes improve crop productivity and provide environmental benefits. Scientifica 2019;2019(1):9106395; CrossRef
53. Lubna, Asaf S, Khan AL, Waqas M, Kang SM, Hamayun M, et al. Growth-promoting bioactivities of Bipolaris sp. CSL-1 isolated from Cannabis sativa suggest a distinctive role in modifying host plant phenotypic plasticity and functions. Acta Physiol Plant 2019;41:1–6; CrossRef
54. Arora P, Tabssum R, Gupta AP, Kumar S, Gupta S. Optimization of indole acetic acid produced by plant growth promoting fungus, aided by response surface methodology. Heliyon 2024;10(14):e34356; CrossRef
55. Silva Santos SD, Silva AAD, Polonio JC, Polli AD, Orlandelli RC, Oliveira JADS, et al. Influence of plant growth-promoting endophytes Colletotrichum siamense and Diaporthe masirevici on tomato plants (Lycopersicon esculentum Mill.). Mycology 2022;13(4):257–27; CrossRef
56. Myo EM, Ge B, Ma J, Cui H, Liu B, Shi L, et al. Indole-3-acetic acid production by Streptomyces fradiae NKZ-259 and its formulation to enhance plant growth. BMC Microbiol 2019;19:1–14; CrossRef
SUPPLEMENTARY MATERIAL
The supplementary material can be accessed at the link here: [https://jabonline.in/admin/php/uploadss/1322_pdf.pdf]